THERYA NOTES 2026, Vol. 7:151-156
Record of tropical boletoid fungi consumption
by the Yucatan brown brocket (Mazama pandora)
in the tropical rainforest of Quintana Roo, Mexico
Registro del consumo de hongos boletoides tropicales
por el temazate café (Mazama pandora)
en la selva tropical de Quintana Roo, México
Romeo Alejandro Sánchez-Zavalegui1, Juan Andrés Estrella-Cach2*, María Marcela Cimé-Ruiz3,
Ismael Pat-Aké2, Javier Isaac de La Fuente4, y Gerardo Sánchez-Rojas5
1Centro de Estudios de Desarrollo Sustentable y Aprovechamiento de la Vida Silvestre (CEDESU), Universidad Autónoma de Campeche. Avenida Agustín Melgar s/n, 24039, San Francisco de Campeche, Campeche, México. E-mail: rasanchz@uacam.mx (RAS-Z).
2Tecnológico Nacional de México (TecNM)-Instituto Tecnológico de la Zona Maya (ITZM), Maestría en Ciencias en Agroecosistemas Sostenibles. Carretera Chetumal–Escárcega km 21.5, Ejido Juan Sarabia, 77965, Othón P. Blanco, Quintana Roo, México. E-mail: andres-estrellaa@hotmail.com (JAE-C), ismael.pa@zonamaya.tecnm.mx (IP-A).
3Universidad Politécnica de Bacalar. Avenida 39 Reg. 12 MZ 325 LT 1 entre calle 56 y 46-A, 77930, Bacalar, Quintana Roo, México. E-mail: maria_cime_ruiz@hotmail.es (MMC-R).
4Colegio de Postgraduados, Edafología. km 36.5, Montecillo, C.P. 56230, Texcoco, Estado de México, México. Email: jdelafuenteitcv@gmail.com (JIL).
5Universidad Autónoma del Estado de Hidalgo, Instituto de Ciencias Básicas e Ingeniería, Área Académica de Biología. Km 4.5 de la Carretera Pachuca Tulancingo s/n Colonia Carboneras, Mineral de la Reforma, Hidalgo 42186. Email: gsanchez@uaeh.edu.mx (GS-R).
*Corresponding author.
Understanding the natural history of species and their feeding habits is essential for their management and conservation. This note documents mycophagy in the brown brocket deer (Mazama pandora) and provides evidence of boletoid mushroom consumption by a tropical cervid. From April to July 2025, 20 passive-sensor camera traps were installed along wildlife trails, near watering holes and in areas covered by fruit trees within the Uaymil Flora and Fauna Protected Area, with a total effort of 1820 camera-trap days. The ingestion of an ectomycorrhizal macromycete by an M. pandora individual was opportunistically documented. To characterize the feeding behavior and mushroom morphology, frames extracted from the camera-trap video were analyzed. An independent event was recorded on July 15, 2025, at 10:54 h, in which an adult individual of M. pandora sniffed, bit, and consumed a boletoid basidioma. The observed characteristics of the pileus, poroid hymenium, and stipe were consistent with species possibly belonging to the genus Neoboletus or Phlebopus, family Boletaceae, order Boletales. This record documents mycophagy by M. pandora in a tropical forest and suggests that, as in temperate forests, cervids may contribute to spore dispersal and ecosystem functioning. These findings highlight the need for further studies on the trophic and ecological importance of macromycetes in the diet of the brown brocket deer.
Key words: Boletales; camera trapping; ectomycorrhizae; mycophagy; tropical rainforest; ungulate.
Comprender la historia natural de las especies y sus hábitos alimenticios es esencial para su manejo y conservación. Esta nota documenta la micofagia del temazate café (Mazama pandora) y aporta evidencia sobre el consumo de hongos boletoides por un cérvido tropical. De abril a julio de 2025 se instalaron 20 cámaras trampa con sensor pasivo en senderos de fauna, abrevaderos y zonas con árboles frutales dentro del Área de Protección de Flora y Fauna Uaymil, con un esfuerzo total de 1,820 días-cámara. De manera fortuita se documentó la ingesta de un macromiceto ectomicorrícico por un individuo de M. pandora. Para caracterizar la conducta alimenticia y la morfología del macromiceto se analizaron fotogramas extraídos del video de la cámara trampa. Se registró un evento independiente el 15 de julio de 2025 a las 10:54 hr, donde un individuo adulto de M. pandora olfateó, mordió y consumió un basidioma boletoide. Las características observadas del píleo, himenio poroide y estípite fueron consistentes con especies del orden Boletales y la familia Boletaceae, posiblemente de los géneros Neoboletus o Phlebopus. Este registro documenta micofagia por M. pandora en selva tropical y sugiere que, como en bosques templados, los cérvidos podrían contribuir a la dispersión de esporas y al funcionamiento del ecosistema. Los resultados evidencian la necesidad de profundizar en la importancia trófica y ecológica de los macromicetos en la dieta del temazate café.
Palabras clave: cámara trampa; ectomicorrizas; micofagia; selva tropical; ungulado.
© 2026 Asociación Mexicana de Mastozoología, www.mastozoologiamexicana.org
DOI: 10.12933/therya_notes-25-245
ISSN 2954-3614
Today, the development of new wildlife monitoring technologies has made it possible to document ecological interactions, including trophic behavior, that were previously difficult to record (Olea et al. 2019). For example, Glebskiy et al. (2024) documented on video the Mexican cottontail (Sylvilagus cunicularius) feeding on eryngo (Eryngium proteiflorum), a feeding habit previously inferred but lacking direct evidence. Similarly, Guimaraes-Silva et al. (2021), using camera traps, documented on video for the first time the predation of quail eggs by Azara agouti (Dasyprocta azarae), a species usually described as granivorous/frugivorous. This record evidenced a trophic interaction that had been previously raised as a behavioral hypothesis.
The feeding habits of wildlife are among the most complex behaviors to document through video recording. One of the most common techniques used in diet research is microhistology (Arellano et al. 2019), which allows the identification of plant species from the epidermis found in feces (Lizcano 2003; Veselovská et al. 2021; Peralta-Pardo et al. 2024). Although this technique has allowed classifying hundreds of plant species consumed by herbivores (Garnick et al. 2018) and has supported hunting management and conservation policies (González and Briones-Salas 2012; Gastelum-Mendoza et al. 2019), it has limitations in detecting ephemeral food types, such as macromycetes, particularly in the case of complex structures such as the pileus and the stipe. Some mycofloristic groups can be identified from the spores, but PCR analysis yields more accurate results in such cases (Pareja et al. 2021).
In cervids, the diet has been widely studied (Serna-Lagunes et al. 2024), mainly due to their importance for hunting and conservation. However, records of macromycete consumption by cervids in Mexico are scarce. Flores-Vázquez (2021) reported mycophagy in Mazama temama, a cervid species living in tropical forests, and this behavior has been also documented in temperate deer, such as reindeer (Rangifer tarandus) and moose (Alces alces) (Launchbaugh and Urness 1992).
This note describes the trophic behavior of a specimen of brown temazate (Mazama pandora), a species listed as Vulnerable according to the IUCN Red List. The videotape of the consumption of a boletoid macromycete by a specimen was recorded fortuitously as part of a larger-scale study. This finding indicates that mycophagy in cervids also occurs in tropical ecosystems, as documented in temperate zones, suggesting potential ecological implications and highlighting the need for further research.
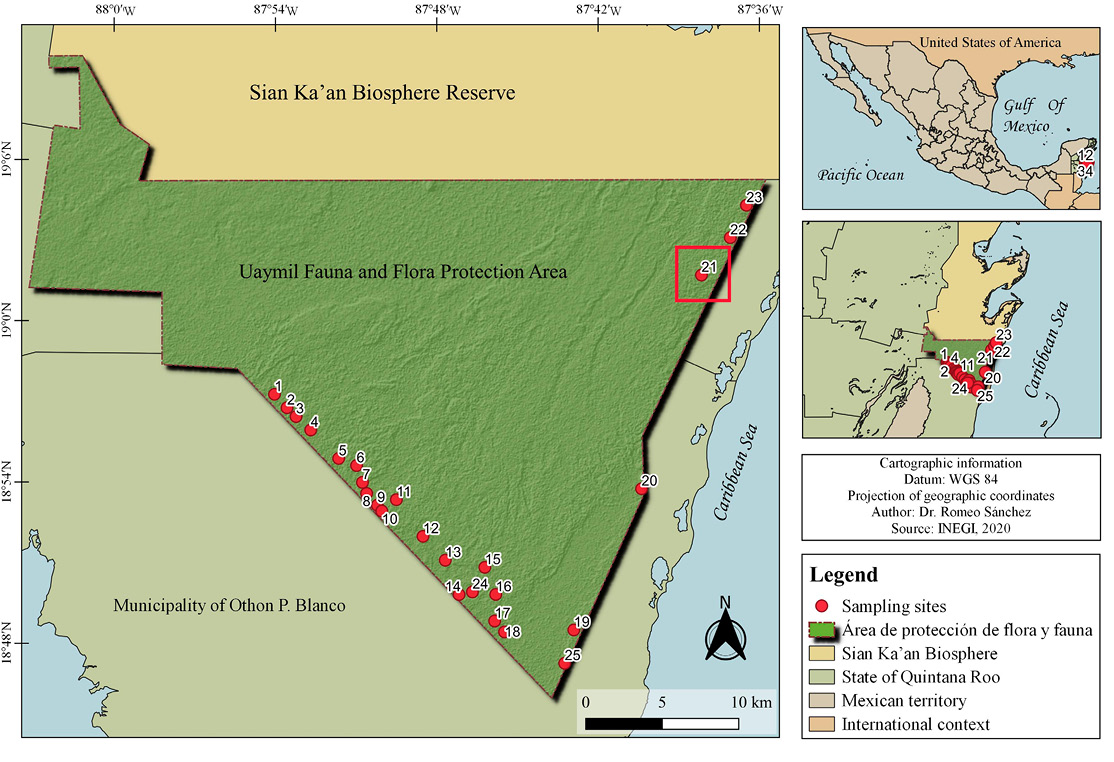
The study area is located in the Uaymil Flora and Fauna Protection Area, Quintana Roo, Mexico (19°1’42.43” N, 87°37’34.41” W), characterized by floodable tropical forest forest of Metopium brownei, Bursera simaruba, and Lysiloma latisiliquum, as well as by mangrove patches of Rhizophora mangle and Conocarpus erectus (Figure 1).
From April to July 2025, a photo-trapping system was installed as part of a larger-scale study focused on wildlife monitoring, following Díaz-Pulido and Payán-Garrido (2012) and Chávez et al. (2013). Twenty Bushnell Prime Low Glow Trail cameras, with a 0.3-second trigger speed, were installed 40–60 cm above the ground along wildlife trails, near watering holes and in areas covered by fruiting trees (Madhusudan and Karanth 2002; Macario-Cueyactle et al. 2019).
The camera traps were in operation for 91 days, accumulating a sampling effort of 1820 camera-days. They were reviewed monthly for record download, and were rotated to new positions at 30-d intervals to cover a larger view of the study area. To reduce bias, exposure to direct sunlight was avoided and the equipment was covered with mud to minimize odor detection. It should be noted that the sampling design was not specifically focused on the study of the diet of M. pandora or the detection of mycophagia; in this context, the reported record corresponds to a fortuitous event recorded as part of the general monitoring of fauna.
An independent event was considered when more than 30 minutes elapsed between two consecutive records of the same specimen. After reviewing the videos, the sequences corresponding to the event reported here were identified; these sequences were processed using the Filmora 12 software to extract the fragments showing the trophic interaction with the macromycete.
Macromycetes were not collected because the record was incidental within a protected natural area and due to the ephemeral nature of fruiting bodies, which were not observed during subsequent field inspections. The consumed macromycetes recorded on video were identified based on the macroscopic characters visible on the video, following the taxonomic keys of Singer et al. (1990) and de la Fuente et al. (2018).
During the sampling period, a mycophagy event was recorded on July 15, 2025 at 10:54 hr at coordinates 19°1’42.43” N, 87°37’34.41” W by the camera trap installed in station 21 (Figure 1). The record occurred during the dry season; however, the area is characterized by high humidity and precipitation due to the relative proximity of the site to the coastline (~3.5 km).
The record corresponded to an adult individual of M. pandora, identified based on its size, morphology, color, distribution, and presence in the region (Sandoval et al. 2023). The visible absence of antlers and the presence of a companion juvenile suggests that it was a reproductively mature female. During the event, the individual sniffed and subsequently consumed a boletoid macromycete (Figure 2a-h). Likewise, traces of substrate removal suggesting the search for other fruiting bodies were identified in the vicinity.
The frames recorded a brown-ochre pileus, a robust stipe and a yellow poroid hymenium (Figure 2c-d), consistent with species in the order Boletales. Although it was not possible to make a specific taxonomic identification, the observed features are compatible with taxa reported for the region, such as Neoboletus antillanus or Phlebopus brasiliensis (de la Fuente et al. 2018; Gelardi et al. 2019).
Throughout the study, M. pandora was recorded on several occasions at different photo trapping stations, suggesting a relatively constant presence in the study area. In contrast, fruiting bodies of boletoid fungi were recorded only occasionally, mainly associated with periods of higher environmental humidity. This difference could indicate that the recorded mycophagia event is an opportunistic feeding habit favored by the temporal availability of the fungal resource.
It should be noted that studies addressing the presence and abundance of mammals in the state of Quintana Roo are still limited. According to Sosa-Escalante et al. (2013) and Sánchez-Zavalegui et al. (2024) there are at least seven published works referring to the fauna of the state, of which three correspond to technical reports developed in protected natural areas. Likewise, the information available specifically for the study area is still scarce.
Macromycete consumption by Mazama pandora is consistent with documented mycophagy patterns of cervids inhabiting temperate regions, where species of the orders Boletales and Amanitales are recurrent components of the diet (Launchbaugh and Urness 1992; Elliott et al. 2022). The record reported herein documents that mycophagy in ungulates is not unique to temperate ecosystems.
Boletoid fungi play central ectomycorrhizal functions in forest nutrient dynamics, and their spores can remain viable after gastrointestinal transit, suggesting a potential role of herbivores in fungus dispersal (Elliott et al. 2022). In tropical ecosystems, this type of ecological interaction has been poorly documented.
In tropical environments, macromycete availability is usually associated with rainy seasons and the phenology of host tree species, so opportunistic fungi consumption could be a nutritional supplementation strategy in periods of high humidity and fungus abundance (Cadotte et al. 2021; Elliott et al. 2022). Similarly, various Boletales species form ectomycorrhizal associations with dominant trees in various forest types, favoring the resistance of their hosts against diseases and environmental stress (Ediriweera et al. 2020).
The existence of an ecological interaction homologous to the one described here has been documented in tropical forests, where macromycete genera such as Russula, Amanita, and Xerocomus establish mycorrhizal symbiosis with trees of the genus Coccoloba (Corrales et al. 2022; Gelardi et al. 2023). The presence of these fungi in tropical environments could facilitate trophic interactions with herbivorous mammals. In this sense, the present record suggests that tropical fungus species may be a food resource for cervids, similarly to their documented role in temperate ecosystems.
The recurrent presence of Mazama pandora in different photo-trapping stations, together with the occasional availability of fruiting bodies during high-humidity periods, suggests that mycophagy could be part of under-recorded trophic interactions in tropical ecosystems. The scarce documentation of this type of event is probably related to methodological limitations and the ephemeral nature of macromycetes.
Audio-visual documentation contributes to mitigate the limitations of indirect dietary analysis methods and provides direct information on poorly recorded trophic interactions. It is recommended to involve molecular analysis of feces and systematic fungus sampling to assess the trophic importance of macromycetes in the diet of M. pandora and its ecological implications related to fungus dispersal and forest dynamics.
Acknowledgements
We thank the master’s peers that assisted in fieldwork. The use of Chat Gpt artificial intelligence (AI) during the manuscript preparation and correction stages is noted; therefore, the authors assume responsibility for the content of this note, noting that all AI-generated content was assessed and edited from the autors’ original ideas.
Literature cited
Arellano, i., et al. 2019. Modificación de la técnica microhistológica. Archivos de Zootecnia 68:164–166.
Cadotte, M., et al. 2021. Mycophagy of white-tailed deer (Odocoileus virginianus Zimmermann) in the boreal forest. Forests 9:1247.
Chávez, c., et al. 2013. Manual de fototrampeo para el estudio de fauna silvestre. Alianza WWF–Telcel, Universidad Nacional Autónoma de México, México.
Corrales, A., et al. 2022. Diversity and distribution of tropical ectomycorrhizal fungi. Mycologia 114: 919–933.
De La Fuente, J. I., et al. 2018. Some interesting Boletales (Basidiomycota) from Quintana Roo, Mexico. Scientia Fungorum 48:77–86.
Díaz-Pulido, A. and E. Payán Garrido. 2012. Manual de fototrampeo: una herramienta de investigación para la conservación de la biodiversidad en Colombia. Instituto de Investigaciones de Recursos Biológicos Alexander Von Humboldt y Panthera Bogota, Colombia.
Ediriweera, A. N., et al. 2020. Linking ectomycorrhizal mushroom species richness and composition with dominant trees in a tropical seasonal rainforest.Studies in Fungi 5: 471–484.
Elliott, T. F., et al. 2022. Mammalian mycophagy: a global review of ecosystem interactions between mammals and fungi. Fungal Systematics and Evolution 9:99–159.
Flores-Vázquez, R. J. 2021. Hábitos alimenticios del venado temazate rojo (Mazama temama), en San Bartolo Tutotepec, Hidalgo, México. Bachelor’s degree dissertation, Facultad de Ciencias, Universidad Nacional Autónoma de México, Ciudad de México, México.
Garnick, S., et al. 2018. Assessment of animal-based methods used for estimating and monitoring rangeland herbivore diet composition. Rangeland Ecology and Management 71:449–457.
Gastelum-Mendoza, F. I., et al. 2019. Dieta de herbívoros: técnica, importancia e implicaciones en el manejo de fauna silvestre. Agro Productividad 12:17–23.
Gelardi, M., et al. 2019. Neoboletus antillanus sp. nov., first report of a red-pored bolete from the Dominican Republic and insights on the genus Neoboletus. MycoKeys 49:73–97.
Gelardi, M., et al. 2023. Coccoloba-associated xerocomoid boletes (Boletaceae) from the Caribbean and Mexico: Tropicoboletus ruborculus gen. et comb. nov., revision of Xerocomus coccolobae, phylogenetic assessment of Singerocomus guadelupae comb. nov., and type studies of Xerocomus caeruleonigrescens, X. cuneipes, and X. pseudoboletinus var. pini-caribaeae. Mycological Progress 22:29.
Glebskiy, Y., et al. 2024. Consumption of Eryngium proteiflorum by the Mexican cottontail (Sylvilagus cunicularius). Therya Notes 5:24–27.
González, G., and M. Briones-Salas. 2012. Dieta de Odocoileus virginianus en un bosque templado del norte de Oaxaca, México. Revista de Biología Tropical 60:447–457.
Guimaraes-Silva, M. A., et al. 2021. Las cámaras trampa revelan la depredación de nidos artificiales por agutíes de Azara en libertad, Dasyprocta azarae Lichtenstein, 1823, en el centro de Brasil. Ecología Austral 46: 148-150.
Launchbaugh, K. L., and P. J. Urness. 1992. Mushroom consumption by North American cervids. Great Basin Naturalist 52:321–327.
Lizcano, D. J. 2003. Ecology and conservation of the mountain tapir in the central Andes of Colombia. Tapir Conservation 12:11–13.
Macario-Cueyactle, D., et al. 2019. Riqueza y abundancia de mamíferos en un ambiente antropizado en Zongolica, Veracruz. Ecosistemas y Recursos Agropecuarios 6:411–422.
Madhusudan, M. D., and K. Karanth. 2002. Local hunting and the conservation of large mammals in India. Ambio 49-54.
Olea, P. P., et al. 2019. Carrion ecology and management. Springer, New York, USA.
Pareja, J., et al. 2021. Complementarity between microhistological analysis and PCR-capillary electrophoresis in diet analysis of goats and cattle using faecal samples. Animal 15:100145.
Peralta-Pardo, R., et al. 2024. Microhistological references of plants available for ungulates in Sonora, Mexico. Agro Productividad 17:119–130.
Sánchez-Zavalegui, R. A., et al. 2024. Presence of medium-sized mammals in a fragment of tropical forest on the banks of the Bacalar Lagoon, Quintana Roo. Therya Notes 5:172–176.
Sandoval, E., et al. 2023. Cytogenetic, molecular, and morphological characterization of Odocoileus pandora (Merriam, 1901) (Artiodactyla, Cervidae). Canadian Journal of Zoology 11:967–979.
Serna-Lagunes, R., et al. 2024. Dieta de Mazama temama en el Parque Nacional Cañón del Río Blanco. Ecosistemas y Recursos Agropecuarios 11:24–69.
Singer, R., et al. 1990. The Boletinae of Mexico and Central America III. Nova Hedwigia Beihefte 102:1–97.
Sosa-Escalante, E., et al. 2013. Mamíferos terrestres de la península de Yucatán, México: riqueza, endemismo y riesgo. Revista Mexicana de Biodiversidad 84:949–969.
Veselovská, A., et al. 2021. A key to the microhistological determination of plant fragments consumed by Carpathian forest deer. Forests 12:1229.
Associate editor: Jorge Ayala Berdón
Submitted: January 19, 2026; Reviewed: May 27, 2026
Accepted: June 16, 2026; Published on line: July 9, 2026
Figure 1. Study area within the Uaymil Flora and Fauna Protection Area, Quintana Roo, Mexico; detail of the observation area in the red box.
Figure 2. (a) Mazama pandora, side view. b) Individual approaching the macromycetus. c–d) View of the boletoid fungus pileus and poroid hymenium. e–g) Basidiom chewing and swallowing. h) Individual after having consumed the macromycete. The original camera trap footage is featured in an explanatory science communication video hosted on the Centro de Investigación de la Universidad Autónoma de Campeche Facebook page (https://www.facebook.com/reel/1297273865219541).
