THERYA NOTES 2026, Vol. 7:120-126
First record of a Tamandua mexicana (Pilosa: Myrmecophagidae) specimen with six cervical vertebrae
Primer registro de un ejemplar de Tamandua mexicana
(Pilosa: Myrmecophagidae) con seis vértebras cervicales
Oscar Daniel González Santana¹*, Samantha Jardon Xicotencatl² y Itze Linda Monroy Hernández¹
¹Área de Fauna Silvestre, Sección de Ciencias Pecuarias, Facultad de Estudios Superiores Cuautitlán. Carretera Cuautitlán–Teoloyucan km 2.5, C.P. 54714, Cuautitlán Izcalli, Estado de México, México. Email: oscar.gonzalez@cuautitlan.unam.mx (ODGS), monroyitze@gmail.com (ILMH)
²Área de Anatomía Veterinaria, Departamento de Ciencias Biológicas, Facultad de Estudios Superiores Cuautitlán. Carretera Cuautitlán–Teoloyucan km 2.5, C.P. 54714, Cuautitlán Izcalli, Estado de México, México. Email: doctora.jardon@cuautitlan.unam.mx (SJX)
*Autor de correspondencia. Teléfono: +52 55 1646 2534
Mammals generally exhibit seven cervical vertebrae, therefore numerical variations are rare and, in most cases, associated with developmental abnormalities. This study reports the first confirmed case of a Tamandua mexicana specimen exhibiting only six cervical vertebrae, providing novel morphological and radiological insights into the family Myrmecophagidae. A juvenile T. mexicana specimen, housed in the Museum Collection of the Facultad de Estudios Superiores Cuautitlán (registry DGVS-CC-344-Mex/24), was examined prior to preservation using the phenolic glycerin technique. Morphometric data and digital radiographs were obtained, and the identification of cervical vertebrae was performed using anatomical criteria established by the Nómina Anatomica Veterinaria. Radiographic examination revealed six cervical vertebrae (C1–C6) with a direct transition to the first thoracic vertebra (T1), without evidence of transitional elements, anomalous fusions, or radiographic signs of trauma or pathological changes. The persistence of open growth plates in the long bones indicated active epiphyseal growth, confirming the juvenile status of the specimen.This case represents the first documented occurrence of cervical numerical variation within Myrmecophagidae, supporting the hypothesis that species with slow metabolic rates may display a higher degree of axial skeletal flexibility. The close phylogenetic relationship between T. mexicana and sloths strengthens this association. A systematic survey of osteological and preserved xenarthran specimens, as well as live individuals, is recommended to determine the frequency and origins of this condition.
Keywords: Comparative anatomy; axial morphology; northern tamandua; Xenarthra; cervical variation
Generalmente los mamíferos presentan siete vértebras cervicales, por lo que las variaciones en el número de estas son raras y en la mayoría de los casos, están asociadas con alteraciones del desarrollo. En este trabajo se reporta el primer registro documentado de un ejemplar de tamandúa (Tamandua mexicana) con seis vértebras cervicales, aportando datos morfológicos y radiológicos relevantes. Se analizó un ejemplar juvenil de T. mexicana procedente de la Colección Museística de la Facultad de Estudios Superiores Cuautitlán (registro DGVS-CC-344-Mex/24) previo a su conservación por la técnica de glicerina fenolada. Se realizaron mediciones morfométricas y radiográficas digitales empleando criterios anatómicos basados en la Nómina Anatómica Veterinaria para determinar el número y características de las vértebras cervicales. Las radiografías mostraron la presencia de seis vértebras cervicales (CI–CVI) con transición directa a la vértebra torácica I (TI), sin evidencias de vértebras transicionales, fusiones anómalas ni signos radiográficos de patología o trauma. La presencia de fisis en los huesos largos evidenció actividad en las líneas de crecimiento, lo que permitió inferir que el ejemplar era juvenil. Este hallazgo constituye el primer registro de variación numérica cervical en Myrmecophagidae, respaldando la hipótesis de que especies con metabolismo lento pueden presentar flexibilidad numérica en el esqueleto axial. La cercanía filogenética de T. mexicana con los perezosos refuerza esta asociación. Se sugiere la revisión sistemática de colecciones y ejemplares preservados de xenartros, así como en ejemplares vivos, para evaluar la frecuencia y causas de esta condición.
Palabras clave: Anatomía comparada; morfología axial; tamandúa norteño; Xenarthra; variaciones cervicales
© 2026 Asociación Mexicana de Mastozoología, www.mastozoologiamexicana.org
Las vértebras cervicales constituyen la base ósea de la región cervical de la columna vertebral de los mamíferos. Esta región desempeña un papel fundamental en el soporte del cráneo, la protección de la médula espinal cervical y la facilitación de los movimientos de la cabeza y el cuello, adaptándose al estilo de vida de cada especie y contribuyendo a su supervivencia en distintos entornos (Singh 2017; Böhmer et al. 2018). Aunque las vértebras cervicales presentan variaciones morfológicas y funciona-les entre especies, los mamíferos muestran una notable constancia en su número, con siete vértebras cervicales en la mayoría de las especies descritas, a diferencia de otros vertebrados como aves y reptiles (Galis 1999). Esta constancia se mantiene a pesar del amplio rango de adaptaciones funcionales, ecológicas y de tamaño corporal presentes en los mamíferos, que abarca desde especies de cuellos muy largos como las jirafas hasta especies con cuellos muy reducidos como las ballenas (Galis 1999). Este patrón ha sido de gran interés en la comunidad científica, ya que durante largo tiempo se desconocieron las causas y consecuencias de la variación en el número de vértebras cervicales. Actualmente, se reconoce que dicha constancia se asocia principalmente a restricciones evolutivas deri-vadas del desarrollo embrionario, así como a los riesgos funcionales y selectivos vinculados con mutaciones que alteran este número (Galis 1999; Buchholtz et al. 2012; Buchholtz 2014; Böhmer et al. 2018).
La consistencia de siete vértebras cervicales en los mamíferos puede interpretarse como el resultado de una evolución conservadora, en la cual cualquier desviación significativa podría comprometer la viabilidad del organismo (Böhmer et al. 2018). Por ejemplo, estudios recientes han demostrado que las alteraciones en el número de vértebras cervicales en mamíferos con metabolismo considerado “normal” se asocian con un incremento significativo en la incidencia de defectos congénitos, mutaciones y tasas de mortalidad en estadios tempranos del desarrollo (Varela-Lasheras et al. 2011; Bhatlekar et al. 2018; Böhmer et al. 2018). Esto se debe a que la segmentación de las vértebras cervicales está estrechamente vinculada a la expresión de los genes Hox, los cuales regulan procesos clave del desarrollo del mesodermo y otros sistemas embrionarios (Deschamps y Duboule 2007; Bhatlekar et al. 2018; Böhmer et al. 2018).
Dentro de los mamíferos solamente tres géneros difieren de esta generalidad: los perezosos (Bradypus y Choloepus) y los manatíes (Trichechus). En especies del género Bradypus, el número de vértebras cervicales se incrementa entre 8-10, mientras que en Choloepus se reduce entre 5–7 (Buchholtz y Stepien 2009). Por su parte, los manatíes presentan de manera constante seis vértebras cervicales (Buchholtz et al. 2007). Estas variaciones representan desviaciones significativas del patrón común y plantean interrogantes sobre los factores evolutivos y funcionales que las han permitido. Estas excepciones se atribuyen a una combinación de presiones selectivas relajadas y adaptaciones funcionales específicas. En el caso de los perezosos, su metabolismo lento y su baja tasa reproductiva podrían haber reducido la presión selectiva en contra de mutaciones que afectan el número de vértebras cervicales (Buchholtz y Stepien 2009). Ambos géneros de perezosos desarrollaron soluciones anatómicas distintas para el soporte del cráneo durante la suspensión corporal, favoreciendo su estilo de vida arborícola lento y críptico. En el género Bradypus, un mayor número de vértebras cervicales se asocia con un rango ampliado de movimiento del cuello, que puede alcanzar hasta 270°, permitiendo la exploración del entorno en posición invertida sin necesidad de desplazar el cuerpo (Miller 1935; Marten et al. 2023). En contraste, el género Choloepus presenta un cuello más corto, con musculatura cervical robusta y una clavícula desarrollada, lo que limita la flexibilidad pero facilita el mantenimiento de una posición extendida para alcanzar el alimento sin movilizar el cuerpo (Miller 1935). Por otro lado, en los manatíes, la transición evolutiva hacia un medio acuático redujo las restricciones mecánicas y funcionales asociadas a la región cervical. Una dieta herbívora y la ausencia de depredadores naturales significativos en su hábitat facilitaron la reducción de la región cervical sin consecuencias adaptativas negativas (Varela-Lasheras et al. 2011).
Los mamíferos de metabolismo y estilos de vida lentos pueden presentan una mayor incidencia de variaciones en el número de vértebras cervicales ya que una tasa metabólica baja se asocia con menores niveles de daño oxidativo al ADN, una reducción en la tasa de mutaciones somáticas y, en consecuencia, con una menor incidencia de efectos pleiotrópicos adversos tales como neoplasias y alteraciones del sistema hematopoyético, lo que disminuye los costos selectivos asociados a desviaciones en el número de vértebras cervicales (Speakman et al. 2002; Hulbert et al. 2007; Bhatlekar et al. 2018; Galis et al. 2022). En este contexto, se podría esperar que especies taxonómicamente cercanas a los perezosos y con un metabolismo relativamente bajo, puedan presentar variaciones en el número de vértebras cervicales. En el presente estudio, se reporta por primera vez una reducción en la cantidad de vértebras cervicales y lumbares en la especie Tamandua mexicana.
El tamandúa norteño o también llamado brazo fuerte (T. mexicana) es una especie de mamífero del orden Pilosa, que comparte con los perezosos (Gibb et al. 2016). Se distribuye desde México hasta el norte de Sudamérica incluyendo Guatemala, Belice, Honduras, El Salvador, Costa Rica, Nicaragua, Panamá, Ecuador, Colombia y Venezuela (Cuarón 2005). Puede ser de hábitos tanto diurnos como nocturnos y se desplaza a nivel del suelo o entre las ramas de los árboles. Los individuos son solitarios a excepción de las hembras que pueden estar acompañadas de su cría (Navarrete y Ortega 2011). En vida libre, su dieta se compone principalmente de hormigas, termitas, meliponas, larvas de escarabajos y de algunos frutos (Montgomery 1985a, b; Brown 2011). Esta especie, generalmente presenta siete vértebras cervicales, de diecisiete a dieciocho vértebras torácicas, tres vértebras lumbares, cinco vertebras sacras fusionadas y un número variable de vértebras caudales (Navarrete y Ortega 2011).
El presente trabajo se realizó en el Laboratorio 4 “Morfología Veterinaria y Biología Celular” de la Unidad de Investigación Multidisciplinaria de la Facultad de Estudios Superiores (FES) Cuautitlán de la Universidad Nacional Autónoma de México (UNAM), en México.
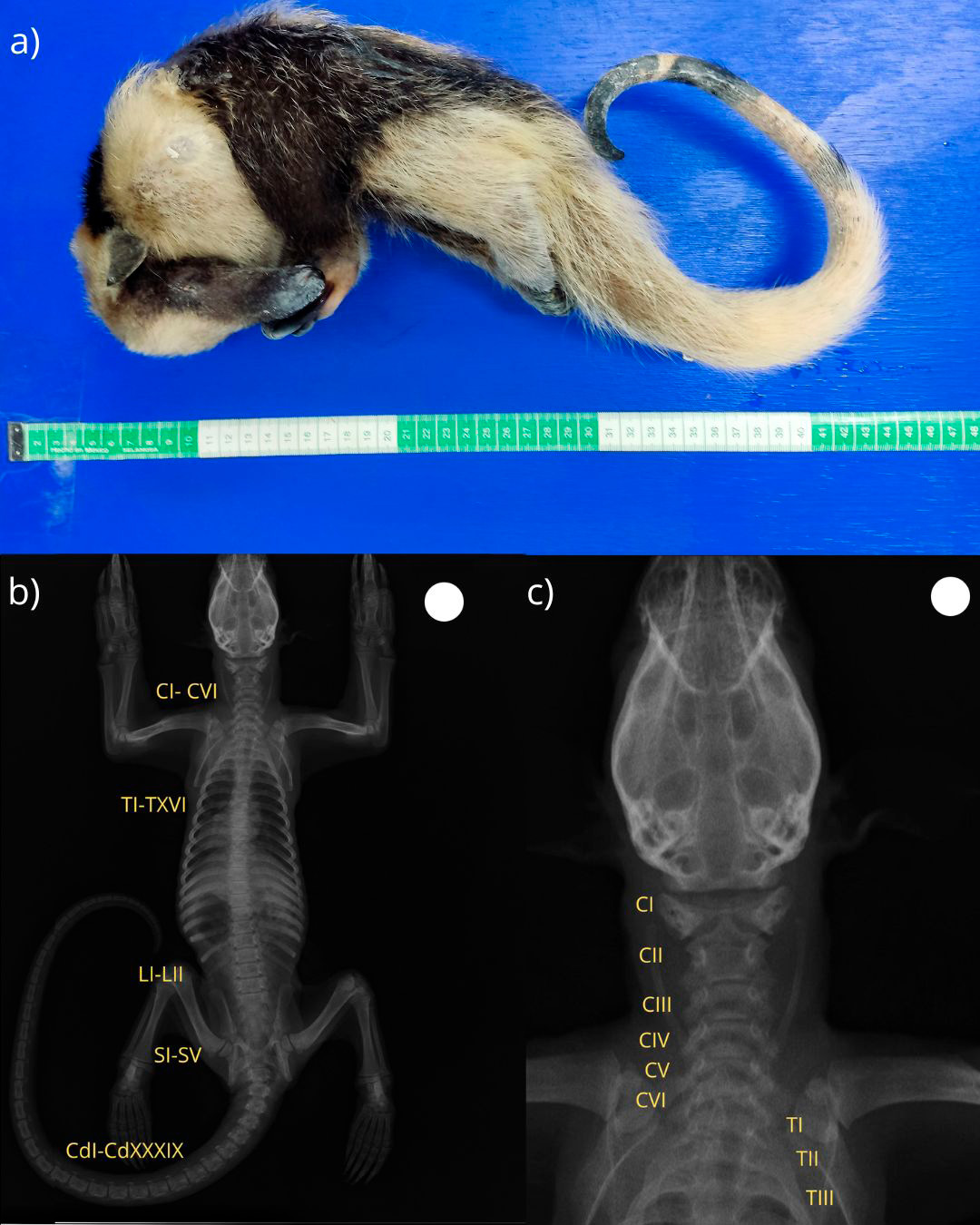
Se utilizó el cadáver de un ejemplar juvenil (Figura 1) de tamandúa (Tamandua mexicana), el cual forma parte de la Colección Museística para la Docencia, la Investigación y la Difusión del Conocimiento de la Fauna Silvestre de la FES Cuautitlán, registrada ante la Secretaría de Medio Ambiente y Recursos Naturales (SEMARNAT) bajo el número DGVS-CC-344-Mex/24. El ejemplar se encontraba conservado en estado de congelación como parte del acervo de la colección museística de la FES Cuautitlán. Al tratarse de material institucional de larga data, los registros históricos disponibles no incluyen información detallada sobre el sitio de procedencia, año de colecta, tipo de vegetación ni condiciones ambientales originales.
El hallazgo objeto de estudio fue identificado previo al procesamiento del ejemplar mediante la técnica de glicerina fenolada, la cual consiste en conservar los tejidos blandos de vertebrados mediante una solución compuesta por glicerina y fenol en proporción 9:1. Posterior al descongelamiento, y como parte de los procedimientos de rutina previos a la preservación, se realizó la toma de medidas morfométricas utilizando cinta métrica y báscula digital.
Para el estudio radiológico, se empleó un equipo de rayos X portátil Soyee modelo SY-31-90P, con potencia máxima de entrada de 1.0 kVA, alimentación AC 50/60 Hz y filtración total equivalente a 2.0 mm Al, realizando proyecciones latero-lateral y ventro-dorsal. Las imágenes obtenidas fueron procesadas mediante un digitalizador FCR Prima II de Fujifilm, sistema de radiografía computarizada (CR) utilizado para la digitalización y optimización de las placas radiográficas para su análisis morfológico. El análisis se efectuó utilizando el visor de imágenes de Windows y la edición de las mismas se realizó con el programa PowerPoint.
El ejemplar reportado correspondió a un macho juvenil de Tamandua mexicana. Aunque la edad exacta no pudo determinarse, su condición juvenil se infirió por la presencia de fisis abiertas en las epífisis de los huesos largos, con líneas de crecimiento no osificadas, además de una talla corporal reducida (Figura 1a). El largo total del cuerpo fue de 83 cm con un peso de 888 g. El sexo se corroboró mediante la observación directa del pene durante la revisión externa del cadáver.
Durante el análisis de las imágenes digitales de las placas radiográficas, se identificó una variación numérica en la región cervical, consistente en la presencia de seis vértebras cervicales (CI–CVI), en lugar del número habitual de siete descrito para la mayoría de mamíferos incluyendo esta especie. La transición se produjo directamente hacia la primera vértebra torácica (TI), sin evidencias de elementos transicionales o fusiones anómalas. La revisión de otros ejemplares (Figura 2) procesados mediante la técnica de glicerina fenolada no mostró esta condición, lo que indica que se trata de un hallazgo aislado. En la radiografía ventro-dorsal de cuerpo completo (Figura 1b), se aprecian todas las estructuras óseas, la diferenciación de las regiones vertebrales y una alineación conservada de la columna vertebral, sin desplazamientos ni deformaciones evidentes.
En el acercamiento radiográfico de la región cérvico-torácica (Figura 1c), se confirma la presencia de seis vértebras cervicales y la transición directa hacia TI, con una morfología conservada en los cuerpos vertebrales y arcos neurales, y sin alteraciones en la articulación atlanto-occipital ni en las costillas de TI.
El conteo completo de las vértebras reveló la siguiente distribución: cervicales: seis (CI–CVI); torácicas: dieciocho (TI–TXVIII), cada una con costillas bien diferenciadas; lumbares: dos (LI–LII); sacras: cinco (SI–SV), fusionadas formando el sacro; y caudales: treinta y nueve (CdI–CdXXXIX), articuladas en serie y con morfología progresivamente simplificada hacia el extremo distal de la cola.
La distribución de las vértebras en el ejemplar de T. mexicana del presente estudio difiere del patrón descrito para la especie T. mexicana, que generalmente presenta siete vértebras cervicales, diecisiete a dieciocho torácicas y tres lumbares, manteniendo un número similar de sacras y caudales (Navarrete y Ortega 2011). La ausencia de una vértebra cervical, en conjunto con la integridad de la alineación vertebral y la ausencia de signos radiográficos de fractura, luxación o remodelación ósea, sugiere que esta variación numérica del esqueleto axial podría ser de origen congénito y no estar asociada a procesos patológicos o traumáticos detectables mediante radiografía.
El presente hallazgo constituye, hasta donde se tiene conocimiento, el primer registro de una variación en el número de vértebras cervicales en Tamandua mexicana y, en general, en cualquier especie de la familia Myrmecophagidae. La constancia de siete vértebras cervicales en mamíferos es un patrón morfológico altamente conservado, mantenido por fuertes restricciones evolutivas y de desarrollo, estrechamente asociadas a la regulación de los genes Hox y a la segmentación axial durante la embriogénesis (Galis 1999; Deschamps y Duboule 2017; Böhmer et al. 2018). Alteraciones en este patrón suelen relacionarse con defectos congénitos, incremento de la mortalidad neonatal y mayor susceptibilidad a enfermedades, incluyendo ciertos tipos de cáncer y disfunciones neuromusculares (Galis 1999; Bhatlekar et al. 2018). Entre los ejemplos documentados, se incluyen una mayor predisposición a ciertos tipos de cáncer, como el cáncer colorrectal y la leucemia mieloide, trastornos del desarrollo hematopoyético y mesenquimal, así como alteraciones neuromusculares, entre ellas el síndrome del desfiladero torácico (Galis 1999; Buchholtz et al. 2012; Bhatlekar et al. 2018; Galis et al. 2022). Por ello, la aparición de variaciones cervicales en poblaciones naturales es considerada un evento raro y, en la mayoría de los casos, potencialmente deletéreo.
Galis et al. (2022) plantearon la hipótesis de que, en mamíferos, la característica conservadora de presentar siete vértebras cervicales puede ser variable en especies de metabolismo basal lento o con un estilo de vida pausado. En tales casos, la relajación de las presiones selectivas contra mutaciones que afectan la morfología axial, combinada con adaptaciones funcionales específicas, parece permitir la persistencia de estos rasgos en las poblaciones. La estrecha relación filogenética entre T. mexicana y los perezosos sugiere que podrían compartir, al menos parcialmente y a nivel del esqueleto axial, esta plasticidad morfológica.
Adicionalmente, en el ejemplar analizado se documentó variación en el número de vértebras lumbares. El presente estudio constituye el primer reporte de una disminución en el número de vértebras cervicales en T. mexicana, mientras que la variación en el número de vértebras lumbares ha sido previamente documentada. Por ejemplo, Gaudin (1999) describió ejemplares con dos vértebras lumbares, concordantes con el presente reporte, y refirió reportes previos de especímenes con tres vértebras lumbares, condición que también es considerada en el diagnóstico anatómico de la especie por Navarrete y Ortega (2011). A diferencia de las vértebras cervicales, las variaciones en el número de vértebras lumbares no se han asociado a efectos pleiotrópicos vinculados a genes del desarrollo, como los documentados para la región cervical y los genes Hox.
En el ejemplar descrito, no fue posible determinar si la reducción a seis vértebras cervicales se debió a una fusión congénita o a una transformación homeótica de la primera vértebra torácica en cervical, fenómenos documentados en otros mamíferos (Böhmer et al. 2018; Galis et al. 2022). En consecuencia, no es posible establecer si esta variación comprometió la funcionalidad del individuo ni si estuvo relacionada con su causa de muerte. La confirmación del origen de esta anomalía requeriría análisis osteológicos detallados o estudios de imagen avanzada, como tomografía computarizada.
Este hallazgo tiene implicaciones relevantes para la anatomía comparada y la biología del desarrollo de los xenartros. Documentar estas excepciones contribuye a comprender la evolución de la columna vertebral en mamíferos y a identificar los límites de la estabilidad morfológica en el eje cervical. Asimismo, resalta la importancia de las colecciones museísticas y de docencia como repositorios de información biológica única. El estudio minucioso de especímenes alojados en dichas colecciones mediante radiografía, tomografía o análisis osteológico puede revelar variaciones morfológicas raras que, de otro modo, pasarían inadvertidas en estudios de campo.
Se recomienda realizar una búsqueda sistemática de casos similares en colecciones osteológicas y ejemplares preservados de Xenarthra, así como la inclusión de monitoreos radiológicos en individuos vivos con el fin de estimar la frecuencia y variabilidad de esta condición, identificar sus posibles bases genéticas y explorar su relevancia adaptativa. Una limitación del presente estudio fue la imposibilidad de conservar y montar el esqueleto del ejemplar, ya que cuando se detectó la variación, el espécimen ya había sido preservado mediante la técnica de glicerina fenolada. Futuras investigaciones podrían superar esta restricción y aportar datos osteológicos más completos que permitan esclarecer el origen y las implicaciones funcionales de esta rara variación cervical.
Agradecimientos
Agradecemos a C. G. García-Tovar, responsable del Laboratorio 4 “Morfología Veterinaria y Biología Celular”, UNAM, por las facilidades otorgadas. El presente artículo fue desarrollado con apoyo de los proyectos PAPIME PE215725 y PIAPIME 2.11.17.25; y las Cátedras: CI2660 y CI2475. Se extiende el agradecimiento a K. A. Hernández-Ponce, F. E. Juárez-Corral y E. M. Hernández-Mendoza por su apoyo.
Literatura citada
Bhateklar, S., Fields, J. Z., y Boman, B. M. 2018. Role of Hox genes in stem cell differentiation and cancer. Stem Cells International 2018:1–15.
Böhmer, C., et al. 2018. Homeotic transformations reflect departure from the mammalian “rule of seven” cervical vertebrae in sloths: Inferences on the Hox code and morphological modularity of the mammalian neck. BMC Evolutionary Biology 18:1–11.
Brown, Danielle. 2011. Fruit-eating by an obligate insectivore: Palmfruit consumption in wild northen Tamanduas (Tamandua mexicana) in Panama en Revista Edentata 12:63–65
Buchholtz, E. A. 2014. Crossing the frontier: A hypothesis for the origins of meristic constraint in mammalian axial patterning. Zoology 117:64–69.
Buchholtz, E. A., Booth, A. C., y Webbink, K. E. 2007. Vertebral anatomy in the Florida manatee, Trichechus manatus latirostris: A developmental and evolutionary analysis. The Anatomical Record 290:624–637.
Buchholtz, E. A., y Stepien, C. C. 2009. Anatomical transformation in mammals: Developmental origin of aberrant cervical anatomy in tree sloths. Evolution & Development 11:69–79.
Buchholtz, E. A., et al. 2012. Fixed cervical count and the origin of the mammalian diaphragm. Evolution & Development 14:399–411.
Cuarón, A. 2005. Oso Hormiguero. Pp 121-123 in Los Mamíferos Silvestres de México (Ceballos,G. y G. Oliva, Coords). Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, Distrito Federal, México.
Deschamps, J., y Duboule, D. 2017. Embryonic timing, axial stem cells, chromatin dynamics, and the Hox clock. Genes & Development 31:1406–1416.
Galis, F. 1999. Why do almost all mammals have seven cervical vertebrae? Developmental constraints, Hox genes, and cancer. Journal of Experimental Zoology 285:19–26.
Galis, F., Van Dooren, T. J. M., y Van Der Geer, A. A. E. 2022. Breaking the constraint on the number of cervical vertebrae in mammals: On homeotic transformations in lorises and pottos. Evolution & Development 24:196–210.
Gasse, H. 2017. Nómina Anatómica Veterinaria. International Committee on Veterinary Gross Anatomic Nomenclature. Hannover, Germany.
Gaudin, T. J. 1999. The morphology of xenarthrous vertebrae (Mammalia: Xenarthra). Fieldiana: Geology 41:1–38.
Gibb, G. C. et al. 2016. Shotgun mitogenomics provides a reference phylogenetic framework and timescale for living xenarthrans. Molecular Biology and Evolution 33:621–642.
Hulbert, A. J., et al. 2007. Life and death: Metabolic rate, membrane composition, and life span of animals. Physiological Reviews 87:1175–1213.
Marten, L. J., et al. 2023. The functional significance of aberrant cervical counts in sloths: Insights from automated exhaustive analysis of cervical range of motion. Proceedings of the Royal Society B 290:1–11.
Miller, R. A. 1935. Functional adaptations in the forelimb of the sloths. Journal of Mammalogy, 16: 38–51.
Montgomery, G. 1985a. Impact of vermilinguas (Cyclopes, Tamandua: Xenarthra = Edentata) on arboreal ant populations. Pp 351-363 in Evolution and Ecology of Armadillos, Sloths and Vermilinguas (Montgomery, G. Ed.), Smithsonian Institute, Washington D. C., EE.UU.
Montgomery, G. 1985b. Movements, Foragin and Food Habitats of the four extant species of neotropical vermilinguas (Mammalia; Myrmecophagidae). Pp 365-370 in The Evolution and Ecology of Armadillos, Sloths and Vermilinguas (Montgomery, G. Ed.), Smithsonian Institute, Washington D. C., EE.UU.
Navarrete, D., y Ortega, J. 2011. Tamandua mexicana. Mammalian Species 43:56–63.
Singh, B. 2017. Dyce, Sack, and Wensing’s Textbook of Veterinary Anatomy. Elsevier, Missouri, EE.UU.
Speakman, J. R., et al. 2002. Living fast, dying when? The link between aging and energetics. Journal of Nutrition 132:1583S–1597S.
Varela-Lasheras, I., et al. 2011. Breaking evolutionary and pleiotropic constraints in mammals: On sloths, manatees and homeotic mutations. EvoDevo 2: 1–27.
Associate editor: Itandehui Hernández Aguilar
Submitted: August 05, 2025; Reviewed: April 06, 2026
Accepted: May 24, 2026; Published on line: June 5, 2026
DOI: 10.12933/therya_notes-25-241
ISSN 2954-3614
Figura 1. a) Ejemplar juvenil de Tamandua mexicana en el que se documentó la variación en el número de vértebras cervicales y lumbares, previo a su preservación anatómica mediante la técnica de glicerina fenolada. b) Imagen radiográfica digital en proyección ventro-dorsal con alineación ósea conservada y sin evidencias de alteraciones en el resto del esqueleto. c) Imagen radiográfica digital en proyección ventro-dorsal con acercamiento a la región cérvico-torácica del mismo ejemplar, evidenciando la presencia de seis vértebras cervicales (CI-CVI) y la transición directa a la primera vértebra torácica (TI).
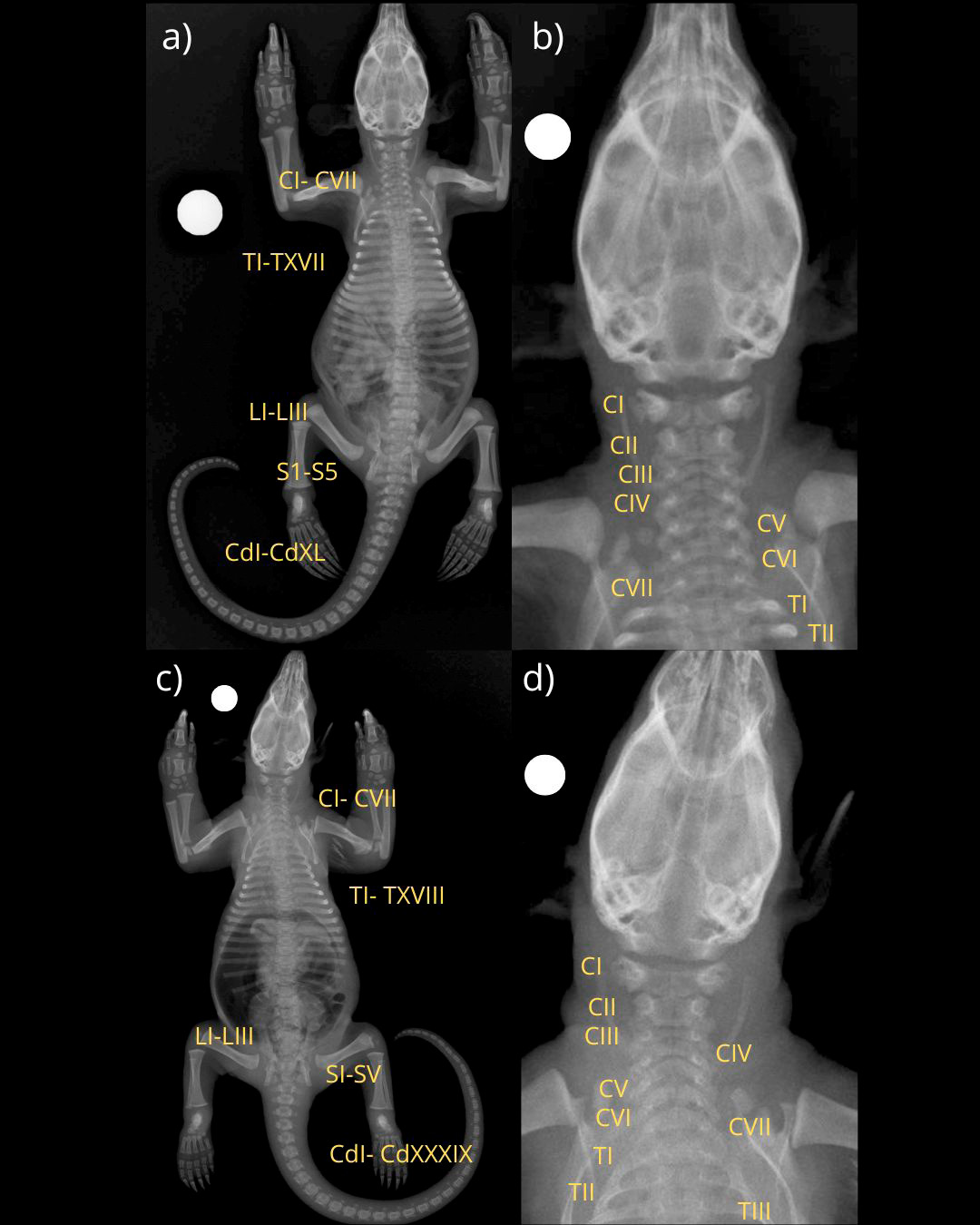
Figura 2. a y c) Imágenes radiográficas digitales en proyección ventro-dorsal de un par de ejemplares juveniles de Tamandua mexicana, con alineación ósea conservada, sin variación numérica en ningún segmento de la columna vertebral y sin evidencias de alteraciones en el resto del esqueleto. b y d) Imágenes radiográficas digitales en proyección ventro-dorsal con acercamiento a la región cérvico-torácica de los mismo ejemplares, evidenciando el patrón cervical normal de esta especie (CI-CVII).