Species coexistence is the ability of multiple species to occupy the same habitat simultaneously and is a dynamic ecological process that is shaped by both intra- and interspecific interactions (Holt 2001; McPeek 2022). Phylogenetically related species often exhibit morphological and behavioral similarities, leading to overlapping resource requirements such as space and food. As a result, competition tends to intensify (Darwin 1859; Blomberg and Garland Jr. 2002). In such cases, particularly among congeneric species, spatial and temporal segregation can serve as key mechanisms to mitigate competitive pressures (Perrin 1980; Castro-Arellano and Lacher Jr. 2009).
Squirrels demonstrate remarkable ecological plasticity and are capable of utilizing both terrestrial and arboreal strata across a broad spectrum of habitats from boreal forests to tropical ecosystems (Best 1995; Lurz et al. 2005). Their adaptability extends even to human-modified landscapes, where they continue to prosper (Koprowski et al. 2017). This flexibility is complemented by complex communication systems, which facilitate frequent intra- and interspecific interactions (McRae and Green 2014; Mazzamuto et al. 2017).
Research on squirrel coexistence has revealed that resource partitioning strategies vary depending on the species involved and the forest structure. These strategies may include differentiation in diet, in vertical space use, and activity patterns (Edwards et al. 1998; Abdullah et al. 2001; Sovie et al. 2019). Such ecological adjustments enable sympatric squirrel species to coexist within shared environments, reducing direct competition and promoting long-term stability in community composition.
Ground-based camera trapping has become a widely used method for studying various aspects of squirrel biology, including abundance, habitat preferences (Shannon et al. 2023), their impact as an invasive species (Beatham et al. 2023), and the expansion of knowledge of the natural history of lesser-known species (Alvarado-Ortíz et al. 2024), across both temperate and tropical regions. More recently, arboreal camera trapping has broadened the scope of mammal monitoring to include both arboreal and terrestrial species (Moore et al. 2021).
In Mexico, the application of arboreal camera trapping has significantly enhanced our understanding of canopy-dwelling mammalian communities (Astiazarán-Azcárraga et al. 2020; Cudney-Valenzuela et al. 2021; Hidalgo-Mihart et al. 2022; García-Casimiro and Santos-Moreno 2024; Rojas-Sánchez et al. 2025). Although squirrel species have been documented in all of these studies, sympatric species were specifically reported only by Cudney-Valenzuela et al. (2021) and Rojas-Sánchez et al. (2025). However, both studies focused on broader mammalian assemblages, and research focused on squirrel ecology remains scarce.
Therefore, studies on the ecology of two sympatric, closely related squirrel species at a local scale present a valuable opportunity to explore spatiotemporal segregation in relation to environmental and vegetation characteristics. Such focused studies can deepen our understanding of niche partitioning and coexistence mechanisms in tropical forest canopies.
In this study, we examined the local frequency and occupancy patterns of two sympatric squirrel species, Sciurus aureogaster and Sciurus deppei, within the limits of the Los Tuxtlas Tropical Biology Station (LTTBS). Our aim was to identify spatiotemporal variations in the use of arboreal habitats in relation to environmental and vegetation characteristics. Specifically, we wanted to quantify how each species utilized vertical and horizontal space over time. We hypothesized that these closely related species would exhibit spatiotemporal segregation as a strategy to reduce competition and facilitate coexistence within the shared landscape.
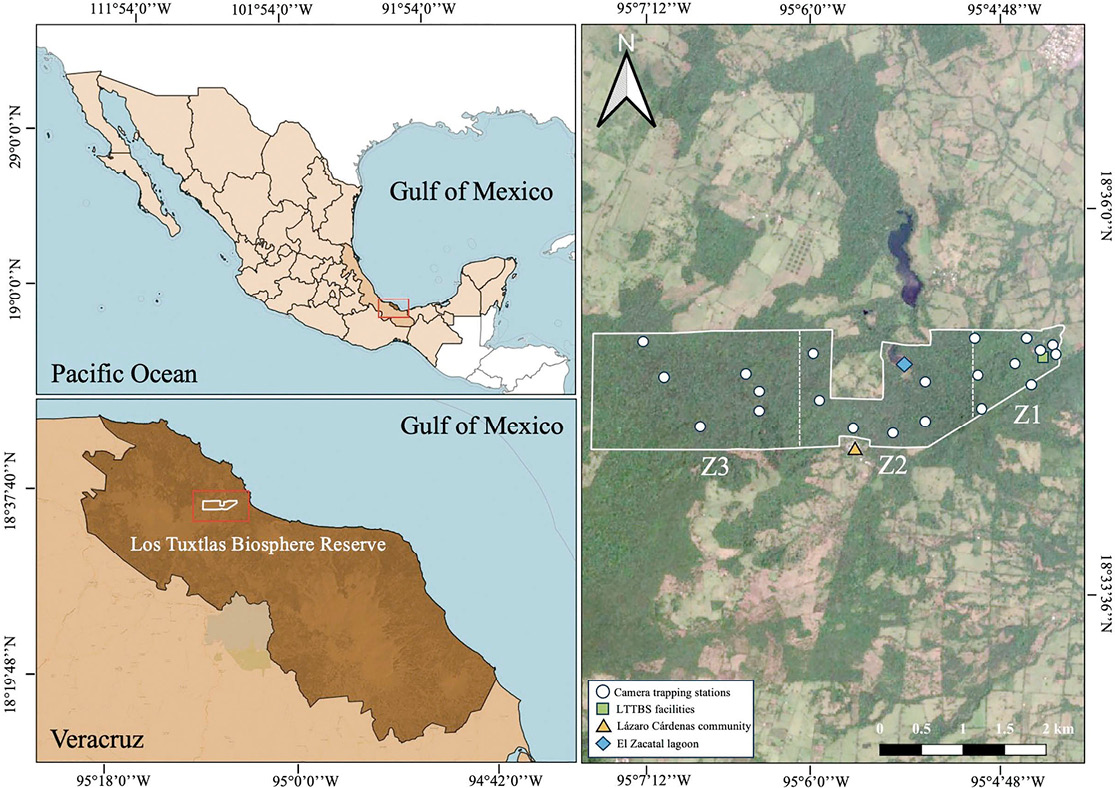
This study was conducted within the Los Tuxtlas Tropical Biology Station (LTTBS), a privately protected area of tropical rainforest owned and operated by the Institute of Biology of the National Autonomous University of Mexico (Coates 2017). The 644-ha reserve is located within the larger Los Tuxtlas Biosphere Reserve (CONANP 2006; Coates 2017; Figure 1). The LTTBS is surrounded by a heterogeneous landscape: the eastern and central portions are bordered by a matrix of pastureland, while the extreme western edge is connected to a remnant of approximately 9,000 hectares of original rainforest, partially surrounding the San Martín Tuxtla volcano (Von Thaden et al. 2020). Notable topographical features include a pronounced slope on the eastern boundary and a seasonal lagoon situated in the central portion of the reserve. Regrettably, on both boundaries (north and south) of the reserve near the central area, the landscape has been modified by the human settlements of the communal lands of Lázaro Cárdenas and the Laguna Escondida.
To monitor arboreal activity, a total of 21 canopy-level camera traps were installed throughout the LTTBS, with one camera per site. These were operated continuously from August 2022 to October 2024. We used a single rope climbing technique for canopy access. The camera models used were the following: 9 - Bushnell Trophy Trail Cameras HD119717, 8 - UOVision Green 30, 3 - Mixmart HC801, and 1 - LTL Acorn 6210. Sites were revisited bi-monthly to replace batteries and memory cards to ensure uninterrupted operation, as well as to restore any inactive units. Cameras were programmed to record 15-second videos per trigger event, without the use of bait. Vegetation was minimally trimmed only when necessary to prevent false triggers.
Site selection was based on three criteria: (1) the geographic location of a pre-established point grid to ensure systematic placement (Figure 1), (2) the presence of canopy bridges as indicators of arboreal transit (Gregory et al. 2017), and (3) the absence of broken branches or colonies of harmful animals such as stinging insects (Moore et al. 2021). Camera heights ranged from 6 to 20.9 meters (mean = 13.80 m, SD = 4.53 m). This was measured in situ using a calibrated measuring tape The mean distance between camera sites was 560.62 meters.
In order to assess environmental variation and its influence on species distribution patterns, the LTTBS reserve was divided into three distinct zones based on topographic and landscape characteristics: Zone 1 (Z1: Cameras 1–9) was located in the eastern portion of the reserve and has the lowest altitude and a pronounced slope that separates it from Zone 2. Zone 2 (Cameras 10–15) was situated in the central portion of the reserve and is in close proximity to the seasonal lagoon “Zacatal”, the Lázaro Cárdenas human settlement, and a larger notable vegetation gap nearby Laguna Escondida in its central-northern boundary. The Zone 3 (Z3: Cameras 16–21) was located in the western portion which is connected to the remnants of original vegetation surrounding the San Martín Tuxtla volcano (Figure 1).
Species were identified using morphological descriptions provided by Coates-Estrada and Estrada (1986), with body size serving as a key distinguishing trait:
Sciurus aureogaster — Typically larger in size, this species exhibits two distinct color morphs: (1) A mixed grey dorsal fur with reddish fur on the ventral side, and (2) A melanistic morph with entirely black fur (Figure 2). Sciurus deppei – Smaller in size it displays a brownish coat with lighter fur on the belly and limbs (Figure 2).
All identifications were validated through comparison with specimens housed in the LTTBS mammal reference collection. Only records with confident species identifications were included in the final database (Figure 2).
A relative abundance index (RAI) was used as a capture frequency proxy and it was estimated as the quotient of the effective records (i.e. number of records after a 24-hour exclusion criterion), and the sampling effort multiplied by 100. The RAI was calculated considering only the effective records to avoid pseudo-replication and to compare the results with previous reports from the area (Flores-Martínez et al. 2022; Rojas-Sánchez et al. 2025). This RAI was calculated for the complete dataset including both species, as well as grouped separately based on the camera location zone (Figure 1). Seasonal variation in capture frequency was evaluated by dividing the complete dataset (2022 to 2024) into two periods: the dry season (March to July), and the wet season (August to February; Soto and Gama 1997). The differences in number of detections between seasons were assessed with a paired t-test in the statistical software R (R Core Team 2023) for both species. A log-transformation was applied only to the effective records of S. aureogaster in the dry season to meet the statistical assumption of normality. No transformation was required for the remaining subsets.
To evaluate the influence of environmental factors on species presence, several variables were measured at each of the 21 camera trap sites: (1) distance to forest edge, (2) distance to the nearest road, (3) distance to human settlements, and (4) camera height. All spatial variables were obtained using Google Earth Pro (Google LLC 2025), while the camera height was recorded directly at each site.
A Principal Component Analysis (PCA) was conducted to explore patterns and relationships among environmental variables and species presence. The analysis was performed using the FactoMineR and factoextra packages in R (Le et al. 2008; Kassambara and Mundt 2020) and visualized with ggplot2 (Wickham 2016). Additionally, a seasonal component was incorporated into the analysis to account for temporal variation in species detection.
A total of 594 species-level records were obtained for both squirrel species across a sampling effort of 5,147 camera trap days (Figure 2). Site Z1 yielded the highest number of records and detection frequency for both species, followed by Z3 and Z2 (Table 1). The total number of records for S. aureogaster was 278, of which 166 were effective records, and had an overall RAI of 3.23. A decreasing gradient in the number of records and RAI for S. aureogaster was observed from Z1 to Z3 (Table 1) and was detected by 6 of the 9 cameras used at Z1 (66%), 4 of 6 at Z2 (66%), and 1 of 6 at Z3 (16%). In the case of S. deppei, 316 total records were obtained, 182 were effective records with an overall RAI of 3.54. This species showed its highest frequency at Z3, followed by Z1 (Table 1), and it was observed by 5 of 9 cameras deployed at Z1 (55%), 3 of 6 at Z2 (50%), and 5 of 6 at Z3 (83%). Furthermore, both species only co-occurred in 5 of the total 21 cameras (24%), 3 of 9 at Z1 (33%), and 1 of 6 at both, Z2 and Z3 (16%).
Table 1. Total number of records for each squirrel species in the LTTBS. N = total number of records, n = effective records, RAI = Relative Abundance Index with 24-hour exclusion criterion.
|
Species
|
N
|
n
|
RAI
|
Z1
|
RAI in Z1
|
Z2
|
RAI in Z2
|
Z3
|
RAI in Z3
|
|
S. aureogaster
|
278
|
166
|
3.23
|
143
|
2.78
|
21
|
0.41
|
2
|
0.04
|
|
S. deppei
|
316
|
182
|
3.54
|
50
|
0.97
|
15
|
0.29
|
117
|
2.27
|
|
Total
|
594
|
348
|
6.76
|
193
|
3.75
|
36
|
0.70
|
119
|
2.31
|
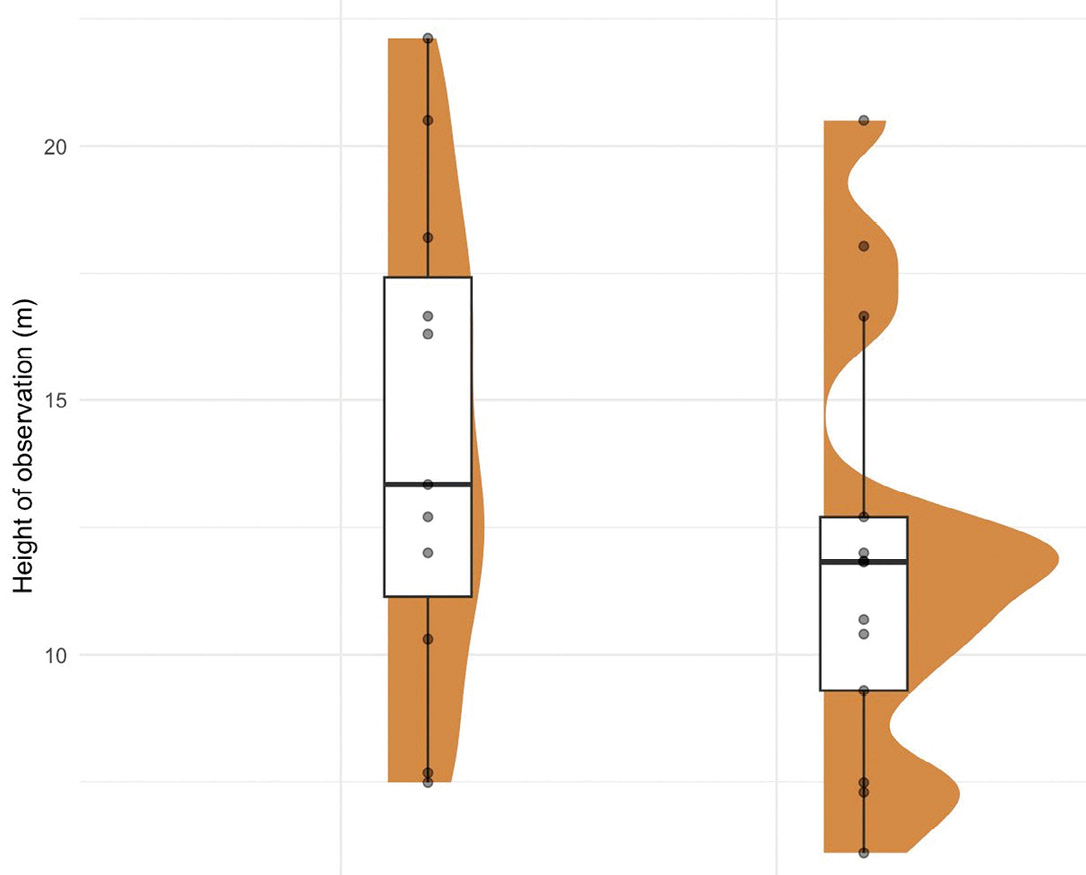
The camera height at which individuals were detected varied between species (Figure 3). Observations of S. aureogaster occurred at heights ranging from 7.49 to 22.1 m (mean = 14.29 m, SD = 4.88), while S. deppei was recorded at lower heights, ranging from 6.1 to 20.5 m (mean = 11.91 m, SD = 4.27). These differences suggest potential vertical stratification in habitat use between the two species.
Regarding seasonal variation, the rainy season yielded the highest number of records for S. aureogaster. In contrast, S. deppei showed only slight differences in detection rates between seasons (Table 2). However, paired t-tests revealed no statistically significant differences in the number of effective records between seasons for either species (S. aureogaster: t = 1.321, P = 0.317, and S. deppei: t = 0.419, P = 0.715 respectively).
The Principal Component Analysis (PCA) revealed that the first principal component (Dim1) accounted for 77.9% of the total variance, while the second principal component (Dim2) explained an additional 18.6%, resulting in a cumulative explained variance of 96.5%. In Dim1, all environmental variables showed significant correlations, however, distance to human settlements had a relatively low contribution to the variance of this component. In contrast, Dim2 was primarily influenced by distance to human settlements, which exhibited the highest contribution and was the only variable with a significant correlation in this dimension (Table 2). These results suggest that while most environmental variables are strongly associated along a shared gradient (Dim1), distance to human settlements may represent a distinct ecological axis influencing species presence independently.
Table 3. PCA associations between S. aureogaster and S. deppei occurrences and the spatial variables based on the R-square values and their corresponding p-value.
|
Variable
|
Dim1
|
p-value
|
Dim2
|
p-value
|
|
Distance to the nearest road
|
0.960
|
1.06 e-5
|
0.160
|
> 0.05
|
|
Distance to forest edge
|
0.918
|
1.76 e-4
|
-0.343
|
> 0.05
|
|
Distance to human settlements
|
0.714
|
2.02 e-2
|
0.688
|
0.02
|
|
Camera height
|
-0.917
|
1.85 e-4
|
0.360
|
> 0.05
|
The PCA biplot (Figure 4) revealed a species-specific segregation along Dim1, while Dim2 primarily reflected temporal variation in species associations with environmental variables. S. aureogaster was predominantly located on the left side of the biplot, indicating a positive association with camera height. Conversely, it showed a negative relationship with distance to forest edges, distance to the nearest road, and distance to human settlements.
This spatial positioning suggests that S. aureogaster tends to occupy strata at higher heights and closer proximity to forest edges, being more associated with anthropogenically influenced zones. The temporal component represented by Dim2 may reflect seasonal shifts in habitat use or detectability patterns for both species. Thus, increased camera height was positively associated with the presence of S. aureogaster throughout the monitoring period, with this relationship being especially pronounced during the rainy season of 2022.
For S. aureogaster, an increase in both distance to human settlements and distance to the nearest road exhibited negative associations, indicating a tolerance for more disturbed habitats. These relationships varied across seasons since distance to the nearest road was more strongly associated with clusters from the dry season of 2023, rainy season of 2023, and dry season of 2024, while distance to human settlements showed a particularly strong negative association during the rainy season of 2024, suggesting increased tolerance to anthropogenic disturbance during this period (Figure 4).
In contrast, S. deppei demonstrated a positive relation-ship with all environmental variables except camera height, indicating a broader sensitivity or avoidance for more open or disturbed areas, as distance to human settlements was positively associated with S. deppei during the rainy season of 2022. Records showed that the increase in distance to an nearest road had a positive effect, especially during the rainy season of 2023 and the dry season of 2023. During both the dry and rainy seasons of 2024, S. deppei was positively associated with distance to forest edges, while camera height showed an inverse relationship, suggesting a preference for lower strata or ground-level activity.
Squirrel species are common and relatively frequent in the Los Tuxtlas Tropical Biological Station (LTTBS), as previously documented (Curiel et al. 1997a, b; Martínez-Gallardo et al. 1997). An earlier arboreal camera-trapping survey conducted by Rojas-Sánchez et al. (2025) identified S. aureogaster as the most frequently observed species (RAI = 2.11), surpassing S. deppei (RAI = 1.68). However, our expanded dataset, which includes an additional 10 months of sampling, reveals a shift in relative abundance: S. deppei was slightly more frequent than S. aureogaster (Table 1).
This shift suggests a rapid increase in detections of S. deppei relative to S. aureogaster, potentially reflecting changes in activity patterns, habitat use, or population dynamics. The notable rise in photo-capture rates of S. deppei in recent months underscores the dynamic nature of species interactions and habitat use within the LTTBS.
In squirrels, reproductive processes are influenced by both resource availability and body mass. However, the duration of resource availability appears to play a more critical role than the mere presence of resources (Hayssen 2008). This may partially explain the observed seasonal and temporal shifts in detection rates, particularly for S. deppei, which showed increased activity during periods associated with greater distance from forest edges and human disturbances.
For instance, we believe that year-to-year phenological variations in food availability within the LTTBS (Dunn et al. 2010) may help explain the observed differences in capture frequency between S. aureogaster and S. deppei. These fluctuations likely influence foraging behavior, movement patterns, and seasonal activity. To validate this hypothesis, future research should incorporate phenological monitoring of food resources from a seasonal perspective, enabling a more nuanced understanding of resource-driven dynamics.
Habitat use between these squirrel species was notably differential, even if both species were present in the three zones within the LTTBS. S. aureogaster exhibited greater occupancy of disturbed areas as in Z1 (Coates-Estrada and Estrada 1986; Koprowski et al. 2017; Curiel et al. 1997a, b). In contrast, S. deppei showed a stronger association with well-conserved habitats, particularly in the western region of the LTTBS (Best 1995; Figure 1). These patterns suggest that habitat specialization may play a role in the coexistence of closely related species.
The ongoing forest fragmentation and landscape transformation at the LTTBS likely create environmental gradients that influence species occupancy and detection rates (Table 1). Previous studies have reported that human-induced habitat fragmentation can increase the abundance of certain vertebrates, including both ground-dwelling and arboreal mammals (Estrada et al. 1984; Rojas-Sánchez et al. 2025). While such changes may reflect habitat degradation, they also offer valuable ecological insights that could provide important information for restoration and management strategies.
Importantly, our findings suggest that S. aureogaster and S. deppei may serve as potential indicators of habitat quality in human-modified landscapes. Their contrasting habitat uses and responses to environmental variables highlight their utility in assessing ecosystem health and directing conservation efforts in tropical forest regions.
Significant correlations between anthropogenic variables —including distance to forest edges, distance to the nearest road, and distance to human settlements— further reinforce the patterns observed in habitat use and species distribution (Table 3; Figure 4). S. aureogaster exhibited positive associations with proximity to these features, aligning with its synanthropic tendencies and documented adaptability to human-modified environments (Hortelano-Moncada et al. 2009; Koprowski et al. 2017). This species’ ability to exploit disturbed habitats may contribute to its persistence in fragmented landscapes.
In contrast, increases in the distance to human disturbance variables such as forest edges, roads and settlements, showed a negative association with S. aureogaster and a positive association with S. deppei. This pattern likely reflects the habitat heterogeneity of the LTTBS, driven by human disturbance, where the most well-conserved forest patches are located at the western portion. Thus, these variables may act as proxies for habitat quality rather than a direct ecological driver, with S. deppei favoring these more pristine areas.
Despite the positive relationship observed between S. deppei and areas with a higher proportion of forest cover, a landscape-scale analysis by Cudney-Valenzuela et al. (2021) reported a negative association between this species and patch size. This apparent contradiction highlights the complexity of habitat selection and suggests that S. deppei may favor smaller, well-connected patches or specific structural features within forested landscapes rather than large, continuous blocks at landscape scale.
To better understand these ecological patterns, an expanded spatial scope within the Los Tuxtlas region is essential. Increasing the range of the study area would allow for a more representative assessment of habitat use and distribution for both species of squirrels, and would enable more robust comparisons with findings from other studies. Such integration is critical for refining conservation strategies and understanding species responses to landscape heterogeneity at multiple scales.
On the other hand, camera height may be considered a non-anthropogenic variable that significantly influences detection, particularly for S. aureogaster, a strictly arboreal species (Koprowski et al. 2017). In contrast, S. deppei frequently descends to the ground for foraging activities (Coates-Estrada and Estrada 1986; Flores-Martínez et al. 2014, 2022), suggesting a spatial segregation that reflects differential vertical space use (Best 1995). Although S. deppei was recorded with high frequency relative to Rojas-Sánchez et al. (2025), higher Relative Abundance Index (RAI) values were consistently observed at ground level (Flores-Martínez et al. 2022), indicating that this species, at least within the LTTBS, exhibits a strong affinity for the understory.
For squirrel species, vertical stratification is a key factor driving niche segregation. Our findings align with patterns of vertical space differentiation reported among sympatric squirrel genera in tropical Malaysian forests (Abdullah et al. 2001). However, in pine-oak forests of Mexico, sympatric sciurids such as S. aureogaster and S. oculatus do not exhibit such vertical segregation (Ramos-Lara and López-González 2017), suggesting that forest type plays a critical role in shaping vertical space use and interspecific interactions.
The lack of significant seasonal variation in records for both squirrel species suggests that seasonality exerts relatively weak influence on their capture frequency within the LTTBS. However, results from the Principal Component Analysis (PCA) revealed a gradual variation in the relationships between squirrels and environmental metrics, underscoring the dynamic nature of habitat requirements (Figure 4). To better understand these patterns, the incorporation of microclimatic data is essential, as abiotic climate-related variables—particularly those differing between canopy and ground-level strata—may play a more nuanced role in shaping ecological responses (Vinod et al. 2023).
Arboreal camera trapping, as employed in our study, has proven to be an effective tool for investigating ecological aspects of canopy-dwelling species (Moore et al. 2021). The ecological responses of squirrels are influenced by multiple dimensions of environmental variation, which interact with morphological, behavioral, and species-specific resource requirements (Edwards et al. 1998). These responses are temporally dynamic, occurring across daily cycles (Sovie et al. 2019), as well as seasonal and annual intervals, as suggested by our findings. Although seasonal differences may not be evident in metrics such as frequency, they may still affect environmental preferences and resource availability. When considered alongside niche partitioning, these subtle shifts could contribute to resource fluctuations that facilitate coexistence between sympatric squirrel species (Edwards et al. 1998).
Temporal multi-scale studies, particularly those involving long-term monitoring, are essential for deepening our understanding of coexistence dynamics among closely related species. Such approaches can help elucidate the environmental drivers behind various ecological patterns such as activity rhythms (Sovie et al. 2019) and are critical for documenting the impacts of climate change and human-induced habitat alterations on arboreal and forest-dwelling mammal species. Integrating these insights can significantly enhance the characterization of squirrels as potential indicator species, offering valuable tools for ecosystem monitoring and conservation planning (Halme et al. 2009).
Acknowledgements
We thank the project “Monitoreo de Mamíferos del Dosel de Los Tuxtlas” from which the results were obtained as part of the main project that served as a requirement for JVR-S in obtaining the M.Sc. degree in the “Posgrado en Ciencias Biológicas, UNAM”. The Consejo Nacional de Humanidades, Ciencias y Tecnologías provided a grant (number 813752) to JVR-S. We also thank the Los Tuxtlas Tropical Biology Station, the Institute of Biology, the NGO Ciencia y Comunidad por la Conservación, and the Idea Wild Organization for the logistical, financial and the equipment provided for the development of this project. Lastly, we thank the reviewers for the suggestions that improved this manuscript.
Literature cited
Abdullah, S.A., N. Yussof-Rashid and A.H. Idris 2001. Niche segregation among three sympatric species of squirrels inhabiting a lowland dipterocarp forest, Peninsular Malaysia. Mammal Study 26: 133-144.
Alvarado-Ortíz, W. J. et al. 2024. Camera-trapping may help to understand the natural history of the deficiently known Glaucomys volans (Rodentia: Sciuridae) in Honduras. Biodiversity 25: 297-299.
Astiazarán-Azcarraga, A., S. Gallina-Tessaro and C. Delfin-Alfonso 2020. Activity patterns of arboreal mammals in a tropical rain forest in México. Therya 11: 225-231.
Beatham, S. E., et al. 2023. A camera trap method for estimating target densities of grey squirrels to inform wildlife management applications. Frontiers in Ecology and Evolution 11:1096321.
Best, T. L. 1995. Sciurus deppei. Mammalian Species 505: 1-5.
Blomberg, S. P., and Garland Jr., T. 2002. Tempo and mode in evolution: phylogenetic inertia, adaptation and comparative methods. Journal of Evolutionary Biology 15: 899-910.
Castro-Arellano, I., and T.E. Lacher Jr. 2009. Temporal niche segregation in two rodent assemblages of subtropical Mexico. Journal of Tropical Ecology 25: 593-603.
Coates, R. I. 2017. La Estación de Biología Tropical Los Tuxtlas desde su fundación. Pp. 13-26 in Avances y Perspectivas en la Investigación de los Bosques Tropicales y sus Alrededores (Reynoso, V. H., R. I. Coates, and M. L. Vázquez-Cruz, eds.). Instituto de Biología, Universidad Nacional Autónoma de México. México City, México.
Coates-Estrada, R., and A. Estrada. 1986. Manual de identificación de campo de los mamíferos de la Estación de Biología Los Tuxtlas. Universidad Nacional Autónoma de México. México City, México.
Comisión Nacional de Áreas Naturales Protegidas. 2006. Programa de Conservación y Manejo Reserva de la Biosfera Los Tuxtlas. Comisión Nacional de Áreas Naturales Protegidas, Mexico City, Mexico.
Cudney-Valenzuela, S., et al. 2021. Does patch quality drive arboreal mammal assemblages in fragmented rainforests?. Perspectives in Ecology and Conservation 19: 61-68.
Curiel, D., V. Sanchez-Cordero and R. Martínez-Gallardo. 1997a. Sciurus aureogaster (ardilla gris). Pp. 622-623 in Historia Natural de Los Tuxtlas (González, E., R. Dirzo and R. Vogt, eds.). Instituto de Biología, Universidad Nacional Autónoma de México, Instituto de Ecología, Universidad Nacional Autónoma de México and Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. México DF, México.
Curiel, D., V. Sánchez-Cordero and R. Martínez-Gallardo. 1997b. Sciurus deppei deppei (ardilla). Pp. 623-624 in Historia Natural de Los Tuxtlas (González, E., R. Dirzo and R. Vogt, eds.). Instituto de Biología, Universidad Nacional Autónoma de México, Instituto de Ecología, Universidad Nacional Autónoma de México y Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. México DF, México.
Darwin, C. 1859. On the origin of species by means of natural selection or the preservation of favoured races in the struggle for life. John Murray. London, U.K.
Dunn, J. C., J. Cristóbal-Azkarate, and J. J. Veà. 2010. Seasonal Variations in the Diet and Feeding Effort of Two Groups of Howlers in Different Sized Forest Fragments. International Journal of Primatology 31:887-903.
García-Casimiro, E., and A. Santos-Moreno. 2024. Activity patterns of an arboreal and semi-arboreal mammal community. Mammal Research 70:35-47.
Google LLC. v 7.3.6.10201. 2025. March 31, 2024. Los Tuxtlas, Veracruz Mexico. 18º 35’ 22.32’’ N, 95º 05’ 48.35’’ W, Eye alt. 9.09 km. Terra Metrics 2025, Airbus 2025. Accessed on 5 February 2025.
Edwars, J.W., D.G.Heckel and D.C. Guynn 1998. Niche Overlap in Sympatric Populations of Fox and Gray Squirrels. The Journal of Wildlife Management 62: 354-363.
Estrada, A., R. Coates-Estrada and C. Vasquez-Yañez. 1984. Observations on Fruiting and Dispersers of Cecropia obtusifolia at Los Tuxtlas, Mexico. Biotropica 16: 315-318.
Flores-Martínez, J. J., et al. 2014. Mamíferos terrestres de la estación de biología tropical de Los Tuxtlas. Revista Digital Universitaria 15:110.
Flores-Martínez, J. J., et al. 2022. Spatiotemporal Coexistence of Mesopredators and Their Prey in a Defaunated Neotropical Rainforest. Tropical Conservation Science 15:1-11.
Halme, P., et al. 2009. Quantifying the indicator power of an indicator species. Conservation Biology 23: 1008-1016.
Hayssen, V. 2008. Reproductive effort in squirrels: ecological, phylogenetic, allometric, and latitudinal patterns. Journal of Mammalogy 89: 582-606.
Hidalgo-Mihart, M. G., et al. 2022. Mammal use of canopy bridges along the Nuevo Xcan-Playa del Carmen highway, Quintana Roo, Mexico. Folia Primatologica 0:1-11.
Holt, R. D. 2001. Species coexistence. Encyclopedia of Biodiversity. Academic Press. Massachusetts, U.S.A.
Hortelano-Moncada, Y., F. Cervantes, and A. Trejo-Ortiz 2009. Mamíferos silvestres de la Reserva Ecológica del Pedregal de San Ángel en Ciudad Universitaria, Universidad Nacional Autónoma de México, México, D.F. Revista Mexicana de Biodiversidad 80: 507-520.
Kassambara, A., and F. Mundt. 2020. factoextra: extract and visualize the results of multivariate data analyses. R package version 1.0.7.
Koprowski, J. L., et al. 2017. Sciurus deppei. Mammalian Species 49: 81-92.
Le, S., J., Josse, and F. Husson. 2008. FactoMineR: An R package for multivariate analysis. Journal of Statistical Software 25: 1-18.
Lurz, P.W., W. Gurnell, and L. Magris 2005. Sciurus vulgaris. Mammalian Species 769: 1-10.
Martínez-Gallardo, R., and V. Sánchez-Cordero. 1997. Historia natural de algunas especies de mamíferos en la región de Los Tuxtlas. Pp. 591-609 in Historia Natural de Los Tuxtlas (González, E., R. Dirzo and R. Vogt, eds.). Instituto de Biología, Universidad Nacional Autónoma de México, Instituto de Ecología, Universidad Nacional Autónoma de México y Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. México DF, México.
Mazzamuto, M. V., et al. 2017. Space invaders: effects of invasive alien Pallas’s squirrel on home range and body mass of native red squirrel. Biological Invasions 19: 1863-1877.
McPeek, M. A. 2022. Coexistence in ecology a mechanistic perspective. Princeton University Press. New Jersey, U.S.A.
Mcrae, T.R., and S.M. Green 2014. Joint tail and vocal alarm signals of gray squirrels (Sciurus carolinensis). Behaviour 151: 1433-1452.
Moore, J., et al. 2021. The potential and practice of arboreal camera trapping. Methods in Ecology and Evolution 12: 1768-1779.
Perrin, M. R. 1980. Ecological strategies of two coexisting rodents. South African Journal of Science 76: 487-491.
R Core team. 2023. R: A language and environment for statistical computing. Vienna, Austria.
Ramos-Lara, N., and C.A. López-González 2017. Niche segregation between Sciurus aureogaster and S. oculatus in a disturbed forest in central Mexico. Journal of Mammalogy 98: 1780-1790.
Rojas-Sánchez, J. V. et al. 2025. Diversity and abundance of the species of arboreal mammals in a tropical rainforest in southeast Mexico. Ecology and Evolution 15: e70812.
Shannon, G., S. Valle, and C.M. Shuttleworth 2023. Capturing red squirrels (Sciurus vulgaris) on camera: A cost-effective approach for monitoring relative abundance and habitat preference. Ecology and Evolution 13: e10536.
Soto, M., and L. Gamma 1997. Climas. Pp. 7-23 in Historia natural de Los Tuxtlas (González-Soriano, E., R. Dirzo, and R. C. Vogt, eds.). Instituto de Biología. Universidad Nacional Autónoma de México. México City, México.
Sovie, A. R, et al. 2019. Ephemeral temporal partitioning may facilitate coexistence in competing species. Animal Behaviour 150: 87-96.
Vinod, N., et al. 2023. Thermal sensitivity across forest vertical profiles: patterns, mechanism, and ecological implications. New Phytologist 237: 22-47.
Von Thaden, J. J., et al. 2020. Dinámica de los cambios en el uso del suelo y cobertura vegetal en la Reserva de la Biosfera Los Tuxtlas (2006-2016). Revista Mexicana de Biodiversidad 91:1-14.
Wickham, H. 2016. ggplot2. Elegant graphics for data analysis. New York, USA.
Associated editor: David Valenzuela Galván.
Submitted: February 24, 2025; Reviewed: July 29, 2025.
Accepted: October 13, 2025; Published on line: December 2, 2025.