THERYA NOTES 2025, Vol. 6: 190-195
Failed predation attempt by a Northern raccoon,
Procyon lotor, on a green iguana, Iguana iguana
Intento fallido de depredación por parte de un mapache,
Procyon lotor, sobre una iguana verde, Iguana iguana
José Manuel Mora1,2*, Nelson Chávez3, Lucía I. López4, and Joel Hause Lewis5
1Carrera de Gestión Ecoturística, Sede Central, Universidad Técnica Nacional (UTN), C. P. 1902-4050. Alajuela, Costa Rica. E-mail: josemora07@gmail.com (JMM).
2Department of Biology and Museum of Vertebrate Biology, Portland State University, C. P. 97207. Portland. Oregon, U.S.A.
3Mistery Jungle, Puerto Viejo, Limón, Costa Rica. E-mail: nelson-11_08@hotmail.com (NC).
4Sede Atenas, Universidad Técnica Nacional. Km 34, Ruta 27, C. P. 7-4013. Balsa de Atenas, Costa Rica. E-mail: llopez@utn.ac.cr (LIL).
5Joe Caribbean Tours, Puerto Viejo, Limón, Costa Rica. E-mail: Joel.lewis89@gmail.com (JHL)
*Corresponding author
The Northern raccoon, Procyon lotor, is distributed from southern Canada to Panama. It occupies a wide range of habitats, including temperate forests, tropical rainforests, dry forests, and disturbed areas, often associated with aquatic environments. The species has also successfully adapted to urban settings. Raccoons are among the most omnivorous mammals known, taking advantage of seasonally abundant food sources. During a guided tour with tourists, we observed a predation attempt by a Northern raccoon on a green Iguana, Iguana iguana, in the Playa Blanca sector of Cahuita National Park, Limón, Costa Rica. This area is part of a well-preserved tropical forest on the Caribbean coast of Costa Rica. The Life Zone is classified as Tropical Moist Forest. On January 7, 2025, at 10:00 hr, we observed a Northern raccoon lunging at a green iguana and biting it on the nape. For approximately 10 secs, the raccoon attempted to subdue the iguana, trying to drag it into the forest. Two seconds later, the iguana broke free and slowly walked toward the edge of the forest and the beach. Raccoons have been reported to prey on mammals up to hare size, but no record exists of them attacking or attempting to prey on animals as large as a green Iguana. However, they are known to prey on turtles reaching 50 cm and 15 kg. This demonstrates that raccoons are capable of preying on relatively large reptiles and mammals.
Key words: Cahuita National Park; Costa Rica; mammals; omnivorous; reptiles; urban adaptation
El mapache, Procyon lotor, se distribuye desde el sur de Canadá hasta Panamá. Ocupa una amplia variedad de hábitats, incluidos bosques húmedos y lluviosos, bosques secos y áreas perturbadas, a menudo asociadas con ambientes acuáticos. La especie también se ha adaptado exitosamente a entornos urbanos. Los mapaches son uno de los mamíferos más omnívoros conocidos y aprovecha fuentes de alimento disponibles estacionalmente. Durante una visita guiada con turistas, observamos un intento de depredación por parte de un mapache sobre una iguana verde, Iguana iguana, en el sector de Playa Blanca, Parque Nacional Cahuita, Limón, Costa Rica. Esta área forma parte de un bosque tropical bien conservado en la costa caribeña de Costa Rica. La zona de vida se clasifica como Bosque Húmedo Tropical. El 7 de enero de 2025, a las 10:00 hr, observamos a un mapache lanzarse sobre una iguana verde y morderla en la nuca. Durante aproximadamente 10 segundos, el mapache intentó someter a la iguana y arrastrarla hacia el bosque. Dos segundos después, la iguana logró liberarse y caminó lentamente hacia el borde del bosque y la playa. Se sabe que el mapache depreda mamíferos de hasta el tamaño de liebres, pero no existen registros de ataques o intentos de depredación sobre animales tan grandes como una iguana verde. Sin embargo, se ha reportado mapaches que depredan tortugas de hasta 50 cm y 15 kg. Esto demuestra que los mapaches son capaces de depredar reptiles y mamíferos relativamente grandes.
Palabras clave: adaptación urbana; Costa Rica; mamíferos; omnívoro; Parque Nacional Cahuita; Reptiles.
© 2025 Asociación Mexicana de Mastozoología, www.mastozoologiamexicana.org
The Northern raccoon, Procyon lotor (Linnaeus 1758), is widely distributed from southern Canada to Panama and occurs at elevations ranging from 0 to 2,800 m (Reid 2009). It has been introduced to several other countries (Stope 2023). In Costa Rica, the Northern raccoon is primarily found in lowland and mid-elevation areas (Mora 2000). It occupies a wide range of habitats, including tropical rainforests (humid and rainy forests), dry forests, and disturbed areas, often associated with aquatic environments (Kays 2009). The species has also successfully adapted to urban settings (Mora 2000).
Raccoons are medium-sized mammals characterized by their distinctive black facial masks and ringed tails (Kays 2009). Their body length ranges from 440 to 620 mm, with a tail length of 190 to 360 mm, and they weigh between 2.7 and 10.4 kg (Kays 2009). This variation in size is largely attributed to their extensive geographic distribution (Helgen and Wilson 2005; Kays 2009).
The raccoon is among the most omnivorous mammals known (Kays 2009; Valenzuela 2014), taking advantage of seasonally abundant food sources (Gerht et al. 2002). The raccoon’s diet is well documented especially in native North America (Gerht 2003). Its diet includes a wide variety of plant materials, such as fruits, nuts, and grains, which constitute most of its year-round diet (Kays 2009). It also consumes a diverse array of invertebrates, including insects and their larvae, terrestrial crabs, crayfish, and occasionally annelids and mollusks (Valenzuela 2014). Aquatic invertebrates, particularly crabs and crayfish, form a significant part of its diet when near water sources, where it also captures fish (Kays 2009). Occasionally, raccoons prey on frogs, lizards, and other small terrestrial vertebrates, though these make up a smaller portion of their diet compared to invertebrates (Kays 2009).
Additionally, raccoons feed on small birds, turtles, mice, squirrels, rabbits, and hares (McAlpine 2011; Valenzuela 2014). They are also known to feed on bats of several genera, such as Myotis in Canada and Indiana, USA (Munson and Keith 1984; McAlpine et al. 2011), and Tadarida in Texas, USA (Winkler and Adams 1972), all of which are relatively small. With its keen sense of smell, the raccoon locates and consumes eggs and nestlings of birds, as well as eggs of turtles, iguanas, and ground-nesting birds (Kays 2009). It also scavenges carcasses of large animals, such as deer, cows, and horses (Valenzuela 2014).
The raccoon is a terrestrial and arboreal species that is primarily nocturnal and generally solitary or found in family groups (Reid and Gómez Zamora 2022). However, individuals may gather in areas with abundant food resources (Kays 2009). Occasionally, raccoons may become active during the day to exploit specific food opportunities, such as foraging during low tide (Kays 2009). In Manuel Antonio National Park on Costa Rica’s Pacific coast, raccoons have adapted their activity patterns to align with visitor hours and days, taking advantage of the increased availability of food provided by human presence (Carrillo and Vaughan 1993).
The green iguana, Iguana iguana (Linnaeus 1758), is the largest lizard species native to Central America. Adult males can reach a total length of up to 2,010 mm (with a standard length of 250–580 mm) and weigh as much as 4 kg (Savage 2002). Females are generally smaller, growing to about 1,440 mm in total length (236–411 mm standard length) and weighing up to 2.9 kg (Savage 2002). This species is easily recognized by its prominent crest of comb-like spines running along the neck, back, and tail, as well as by a large, smooth scale located near the rear margin of the head below the ear opening on the throat (Savage 2002). Adults are typically green—like the juveniles—and often exhibit dark wavy crossbands across the back, bluish shoulder streaks, black abdominal stripes, and dark rings on the tail (Savage 2002). Exceptionally large males may appear uniformly gray, gold, or tan, and during the breeding season, they often develop striking reddish-orange or golden hues along the back (Savage 2002).
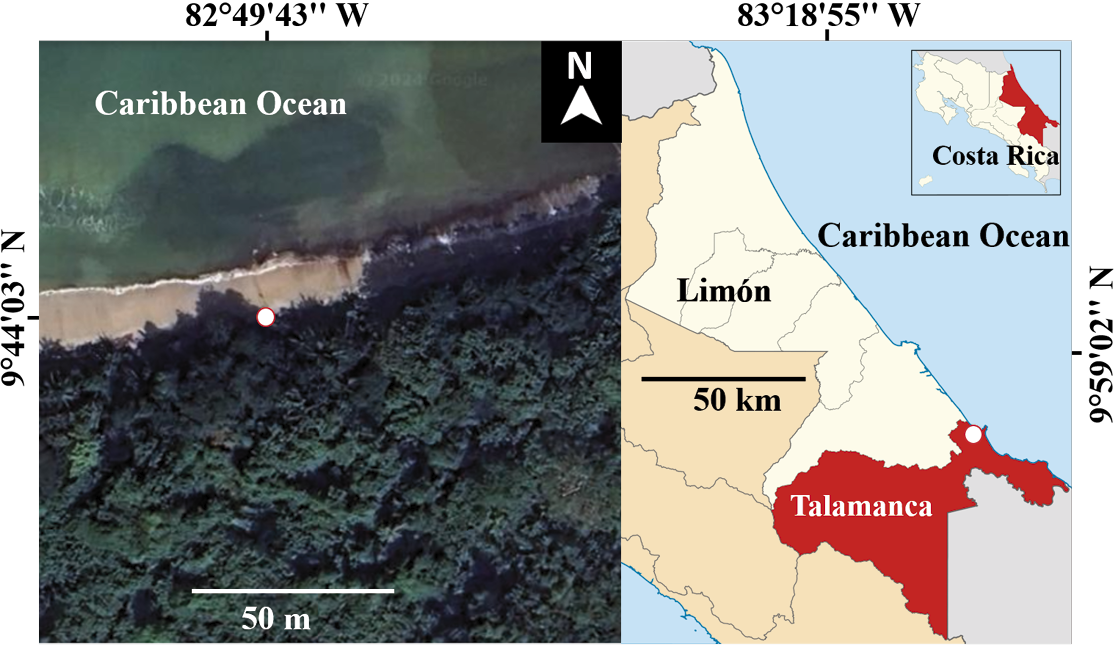
During a guided tour with tourists in Cahuita National Park, we observed a predation attempt by a Northern raccoon on a green iguana. The observation took place in the Playa Blanca sector of the park, located in Limón Province, Costa Rica (9° 44’ 03” N, 82° 49’ 43” W, 7 m; Figure 1). This area is part of a well-preserved tropical forest on the Caribbean coast of Costa Rica (Mora et al. 2020). The Life Zone, according to Holdridge’s system (Holdridge 1967), is classified as a Tropical Moist Forest. Precipitation in this life zone ranges from 1,800 to 4,000 mm annually, with consistently high temperatures and a dry period lasting from 0 to 2 months (Hartshorn 1983). The forest is evergreen and characterized by very high density and typically comprises 4 to 5 distinct strata (Bolaños et al. 2025). The understory is abundant and predominantly composed of evergreen species. The average canopy height ranges between 30 and 40 m, with emergent trees reaching heights of 40 to 50 m (Hartshorn 1983). These canopy trees have wide and elevated crowns, remaining unbranched for the first 25 to 35 m. Their diameters at breast height (DBH) are generally less than 100 cm, and they feature tall, slender, and smooth buttresses (Bolaños et al. 2025).
On January 7, 2025, at 10:00 hr we observed a male green iguana standing in the middle of a trail as several tourists passed by. The tourists paused to watch, with some taking photographs. The iguana displayed its uniform gold to red-orange coloration, typical of adult males during the breeding season (Savage 2002). Only 6 s later, a Northern raccoon lunged at the iguana, biting it on the nape in a manner resembling the hunting behavior of felid carnivores. Upon noticing the approaching raccoon, the iguana raised its body but did not flee.
For approximately 10 s, the raccoon attempted to subdue the iguana, trying to drag it into the forest, where 3 additional raccoons were present, presumably 3 juveniles observing their mother’s hunting behavior. The iguana resisted, struggling to escape toward the opposite side of the trail. At 18 s into the attack, one of the juvenile raccoons approached but retreated when the iguana rolled onto its side in an attempt to free itself. Throughout the encounter, the mother raccoon maintained its grip on the iguana’s nape, occasionally readjusting its bite (Figure 2).
Two seconds later, the iguana broke free and slowly walked toward the edge of the forest and the beach. The raccoon retreated along the trail, while one of the juveniles followed the iguana at a distance of approximately 1 m, sniffing and observing it before returning to the group. The iguana appeared unharmed and exhibited normal behavior. It perched on a log, showing no visible injuries to its nape despite being held in the raccoon’s jaws for several seconds. A video of the predation attempt can be viewed at the following link: https://www.instagram.com/reel/DEi8grzIF4h/?igsh=Y3QxZGN0dWk3azht.
A potential explanation for the observed predation attempt by a raccoon on a green iguana may involve the participation and behavioral development of juvenile individuals within the group. Juvenile raccoons often engage in social learning, acquiring essential foraging and hunting skills through the observation and imitation of conspecifics (Stanton et al. 2024). This process may be part of an extended learning period or “training”, where younger raccoons experiment with different prey and strategies, even when the prey is larger or more challenging to subdue (Daniels et al. 2019; Stanton et al. 2024). Such behavior has been noted in other mammal species where juveniles mimic adults or engage in exploratory behavior that, while not always successful, is crucial for skill development (Thornton and Clutton-Brock 2011).
Another explanation could relate to resource scarcity or changes in local food availability. Raccoons are highly opportunistic and adaptable, and in areas where their usual food sources are reduced—whether due to seasonal variation or human impact—they may broaden their dietary niche and take greater risks in pursuing large or non-traditional prey (Daniels et al. 2019). Nutritional stress or competition for food can increase boldness and lead to novel predatory attempts (Prange et al. 2003; Bateman and Fleming 2012). This may be particularly relevant in disturbed or human-altered environments, where ecosystem balance and food availability fluctuate unpredictably.
Although raccoons have been reported to prey on mammals up to the size of a hare (Valenzuela 2014), there are few documented cases of attacks on larger vertebrates. Notably, a raccoon was observed preying on a juvenile green iguana in Florida, U.S.A., where the species was introduced (Smith et al. 2006). Just a month later, another raccoon attacked a large adult male green iguana in the same location, although the iguana managed to escape (Smith et al. 2006).
Raccoons have also been identified as predators of adult snapping turtles (Chelydra serpentina Linnaeus 1758), a species considered at risk in Canada (Karson et al. 2018). Adult snapping turtles can reach shell lengths of approximately 50 cm and weigh up to 15 kg (Campbell 1998), illustrating that raccoons are capable of preying on relatively large reptiles and mammals. Nevertheless, this represents the first documented case of an attempted, albeit unsuccessful, predation by a raccoon on a green iguana within its native range.
A male green iguana, such as the one observed during the raccoon attack in Cahuita National Park, can exceed 2 m in length (Savage 2002). This considerable size likely enabled the iguana to escape its attacker. Nevertheless, raccoons are clearly capable of preying on smaller iguanas, including females and especially juveniles. Other potential raccoon prey in Cahuita includes basilisks, such as the green basilisk, Basiliscus plumifrons (Cope 1875), and the brown basilisk, Basiliscus basiliscus Linnaeus, 1758, whose maximum male sizes are 900 mm and approximately 600 g, respectively (Savage 2002).
Both iguanas and basilisks are diurnal reptiles (Savage 2002), whereas raccoons are primarily nocturnal (Reid and Gómez Zamora 2022). However, raccoons have been observed shifting their activity patterns toward increased diurnal behavior (Carrillo and Vaughan 1993), potentially increasing the vulnerability of these lizards to predation. The Northern Raccoon is now one of the most frequently observed mammals along the trails of Cahuita National Park at any time of day. In such conditions, raccoons may compete with other predators in the area, including mammals and birds of prey. They are also well-known predators of both continental and marine turtle eggs and ground-nesting birds, particularly when raccoon populations are high (Erickson and Scudder 1947; Stancyk 1982; MacLaren 1992; Kauhala 1996; Rogers and Caro 1998; Engeman et al. 2002, 2005; Schmidt 2003; Beltrán-Beck et al. 2012; Munscher et al. 2012; Kudaktin 2019; Stope 2023). Understanding these dynamics is essential for interpreting raccoon behavior within broader ecological systems.
While it is unclear whether raccoons have been reported in human settlements near Cahuita, their potential presence could influence conservation efforts for vulnerable species. Rather than their feeding habits alone, it is the population density of raccoons that may have the most significant ecological impact, especially in sensitive environments like Cahuita National Park. High raccoon densities can intensify predation pressure on native fauna and disrupt predator-prey dynamics (Riley et al. 1998; Rosatte et al. 2000; Stope 2023). Consequently, management actions must be implemented to protect key species, with local community involvement playing a vital role in ensuring success.
In this context, it is essential to implement regular raccoon population surveys to detect increases in abundance. Where necessary, humane population control measures—such as live trapping and relocation or sterilization programs—should be carried out, particularly in ecologically sensitive areas. Reducing human-related food sources (e.g., unsecured trash, compost, pet food) that artificially sustain raccoon populations is also critical. Waste management should be improved within and around protected areas, including the installation of raccoon-proof bins. Additionally, educating local residents and visitors about the ecological impacts of feeding wildlife is vital. Promoting citizen science initiatives that engage communities in monitoring raccoon activity and reporting predation events can further strengthen conservation efforts.
This study highlights the need for greater attention to the ecological role of raccoons, particularly their predatory impact on vulnerable species such as the green iguana. Understanding raccoon behavior and population dynamics is essential for implementing effective conservation strategies to protect local biodiversity in Cahuita National Park.
Acknowledgements
J. M. Mora acknowledges E. Rivera, Department Head of Ecotourism Management (GEC) at the Central Campus, Universidad Técnica Nacional (UTN), for her academic support. Two anonymous reviewers helped improve an earlier version of this note.
Literature cited
Bateman, P. W., and P. A. Fleming. 2012. Big city life: carnivores in urban environments. Journal of Zoology 287:1–23.
Beltran-Beck, B. F. J. García, and C. Gortazar. 2012. Raccoons in Europe: disease hazards due to the establishment of an invasive species. European Journal of Wildlife Research 58:5–15.
Bolaños, R., V. Watson, and J. Tosi. 2005. Mapa ecológico de Costa Rica (Zonas de Vida), según el sistema de clasificación de zonas de vida del mundo de L. R. Holdridge, Scale 1:750 000. Centro Científico Tropical. San José, Costa Rica.
Campbell, J. A. 1998. Amphibians and reptiles of Northern Guatemala, the Yucatán, and Belize. University of Oklahoma Press, Norman, Oklahoma, U. S. A.
Carrillo, E. and C. Vaughan. 1993. Variación en el comportamiento de Procyon spp. (Carnivora: Procyonidae) por la presencia de turistas en un área silvestre de Costa Rica. Revista de Biología Tropical 41:843–848.
Daniels, S. E., R. E. Fanelli, A. Gilbert, and S. Benson-Amram. 2019. Behavioral flexibility of a generalist carnivore. Animal Cognition 22:387–396.
Engeman, R. M., et al. 2002. An economic analysis of predator removal approaches for protecting marine turtle nests at Hobe Sound National Wildlife Refuge. Ecological Economics 42:469–478.
Engeman, R. M., et al. 2005. Dramatic reduction in predation on marine turtle nests through improved predator monitoring and management. Oryx 39:318–326.
Erickson, A. B., and H. I. Scudder. 1947. The raccoon as a predator on turtles. Journal of Mammalogy 28:406–407.
Gerhrt, S. D. 2003. Raccoon (Procyon lotor) and allies. Pp. 611–663 in Wild mammals of North America: biology, management, and conservation (Feldhamer G. A., B. C. Thompson, and J. A. Chapmanet, eds.). JHU Press, Maryland, U.S.A.
Gerht, S. D., G. F. Huber, and J. A. Ellis. 2002. Long-term population trends of Raccoons in Illinois. Wildlife Society Bulletin 30:457–463.
Hartshorn, G. S. 1983. Plants. Pp. 136–141 in Costa Rican natural history (Janzen, D. H., ed.). University of Chicago Pres. Illinois, U.S.A.
Helgen, K. M. and D. E. Wilson. 2005. A systematic and zoogeographic overview of the raccoons of Mexico and Central America. Pp. 221–236 in Contribuciones Mastozoológicas: en Homenaje a Bernardo Villa (Sánchez-Cordero, V. and R. A. Medellín, eds.). Instituto de Biología e Instituto de Ecología, UNAM, México.
Holdridge, L. R. 1967. Life zone ecology. Tropical Science Center, San José, Costa Rica.
Kauhala, K. 1996. Introduced carnivores in Europe with special reference to central and northern Europe. Wildlife Biology 2:197–204.
Karson, A., S. Y. J. Angoh, and C. M. Davy. 2018. Depredation of gravid freshwater turtles by Raccoons (Procyon lotor). The Canadian Field-Naturalist 132:122–125.
Kays, R. W. 2009. Family Procyonidae (Racoons). Pp. 504–530 in Handbook of the mammals of the world: Vol. 1 Carnivores (D. E. Wilson and R. A. Mittermeier, eds.). Lynx Editions, Barcelona, Spain.
Kudaktin, A. N., and A. V. Romas. 2019. Raccoon in the Russian Black Sea Coast. Biota & Environment 2:88–103.
MacLaren, P. A. 1992. Raccoon (Procyon lotor) depredation on listed and other species of native fauna. Office of Resource Management, Florida Department of Natural Resources. Gainesville, Florida, U.S.A.
McAlpine, D. F., et al. 2011. Consumption of bats (Myotis spp.) by raccoons (Procyon lotor) during an outbreak of white-nose syndrome in New Brunswick, Canada: implications for estimates of bat mortality. The Canadian Field-Naturalist 125:257–260.
Mora, J. M. 2000. Mamíferos silvestres de Costa Rica. Editorial UNED. San José, Costa Rica.
Mora, J. M., L. I. López, and N. Campos. 2020. An adult male Basiliscus plumifrons Cope, 1875 without an arm survive tropical rain forest of Costa Rica. Revista Latinoamericana de Herpetología 3:123–125.
Munscher, E. C., et al. 2012. Decreased nest mortality for the Carolina diamondback terrapin (Malaclemys terrapin centrata) following removal of raccoons (Procyon lotor) from a nesting beach in northeastern Florida. Herpetological Conservation and Biology 7:176–184.
Munson, P. J., and J. H. Keith. 1984. Prehistoric raccoon predation on hibernating Myotis, Wyandotte Cave, Indiana. Journal of Mammalogy 65:152–155.
Prange, S., S. D. Gehrt, and E. P. Wiggers. 2003. Demographic factors contributing to high raccoon densities in urban landscapes. The Journal of Wildlife Management 67:324–333.
Reid, F. A. 2009. A Field Guide to the mammals of Central America and Southeast Mexico. Second edition. Oxford University Press. Oxford, England.
Reid, F., and G. Gómez Zamora. 2022. Pocket guide to the Mammals of Costa Rica. Comstock Publishing Associates. Ithaca, U.S.A.
Riley, S. P. D., J. Hadidad, and D. A. Mansli. 1998. Population density, survival, and rabies in raccoons in an urban national park. Canadian Journal of Science 76:153–164.
Rogers, C. M., and M. J. Caro. 1998. Song sparrows, top carnivores and nest predation: a test of the mesopredator release hypothesis. Oecologia 116:227–233.
Rosatte, R. C. 2000. Management of raccoons (Procyon lotor) in Ontario, Canada: do human intervention and disease have significant impact on raccoon populations? Mammalia 64:369–390.
Savage, J. M. 2002. The amphibians and reptiles of Costa Rica: a herpetofauna between two continents, between two seas. University of Chicago Press. Chicago, Illinois, U.S.A.
Schmidt, K. A. 2003. Nest predation and population declines in Illinois songbirds: a case for mesopredator effects. Conservation Biology 17:1141–1150.
Smith, H. T., et al. 2006. Raccoon predation as a potential limiting factor in the success of the green iguana in southern Florida. Journal of Kansas Herpetology 20:7–8.
Stancyk, S. E. 1982. Nonhuman predator, of sea turtles and their control. Pp. 139-152, in Biology and Conservation of Sea Turtles (Bjorndal, K. A., ed.). Smithsonian Institution Press, Washington, D. C., U.S.A.
Stanton, L. A., et al. 2024. Wild raccoons demonstrate flexibility and individuality in innovative problem-solving. Proceedings of the Royal Society B 291:20240911.
Thornton, A., and T. Clutton-Brock. 2011. Social learning and the development of individual and group behaviour in mammal societies. Philosophical Transactions of the Royal Society B: Biological Sciences 366:978–987.
Winkler, W. G., and D. B. Adams. 1972. Utilization of southwestern bat caves by terrestrial carnivores. American Midland Naturalist 87:191–200.
Valenzuela, D. 2014. Procyon lotor (Linnaeus, 1758) Common Raccoon. Pp. 571–573 in Mammals of Mexico (Ceballos, G., ed.). Johns Hopkins University Press. Baltimore, Maryland, U.S.A.
Associate editor: David Valenzuela-Galván
Submitted: April 04, 2025; Reviewed: October 13, 2025.
Accepted: November 03, 2025; Published on line: December 9, 2025.
DOI: 10.12933/therya_notes-24-220
ISSN 2954-3614
Figure 1. Location (white dot) where a Northern raccoon, Procyon lotor, attempted to prey on a green iguana, Iguana iguana, near the forest edge adjacent to Blanca Beach, Cahuita National Park, in Talamanca County (highlighted in red on the map to the right), Limón Province (outlined in white and shown in red on the map of Costa Rica). The figure was prepared by the author using Google Earth (left section) and Wikipedia under the Creative Commons Attribution-ShareAlike 3.0 license (right section).

Figure 2. A Northern raccoon, Procyon lotor, attempting to subdue an adult male green iguana, Iguana iguana, near the forest edge adjacent to Blanca Beach, Cahuita National Park, Limón Province, Costa Rica.