Mammalian coloration is far from a trivial trait. To date, camouflage appears to be the primary explanation for overall coloration, whereas localized patches of colored fur are generally associated with intraspecific signaling (Caro 2005). Background matching may account for the white pelage, while black-and-white combinations are often associated with aposematism and conspecific signaling. Contrasting coloration can play a significant role in crypsis, mainly through mechanisms such as pattern blending and disruptive coloration, among other functions (Caro 2009). Additionally, Gloger’s ecogeographical rule associates darker coloration with more humid environments. As Delhey (2019) points out, the two most plausible mechanisms underlying this pattern are camouflage and protection against parasites or pathogens, with the latter potentially involving pleiotropic effects on the immune system.
Chromatic disorders in mammals result from either excess (hyperpigmentation or melanism) or a deficiency (hypopigmentation) of melanin (Abreu et al. 2013). Hypopigmentation disorders include albinism, leucism, piebaldism, and hypomelanism (such as flavism, erythrism, and rufism). Albinism is an inherited condition characterized by a complete lack of melanin due to the absence of enzyme tyrosinase. Leucism is a condition characterized by a total or partial loss of pigmentation throughout the body, resulting in a white, whitish, or yellowish appearance. This condition is caused by a recessive gene that inhibits melanin synthesis, although it rarely affects hairless areas, such as the nose, feet, and other exposed skin, and it never alters the pigmentation of the iris (Miller, 2005). As a result, the eyes and/or extremities may retain their original coloration (Fertl and Rosel 2002; Miller 2005; Acevedo and Aguayo 2008). Piebaldism is characterized by localized areas of depigmentation resulting from the absence of melanocytes in affected skin and hair follicles, typically caused by mutations in several genes (Lamoreux et al. 2010). Hypomelanism results in hair varying shades of red or yellow, while eye coloration remains unchanged. This condition is linked to the synthesis of pheomelanin and may be caused by mutations that affect melanin biosynthesis, pigment granule trafficking, or membrane sorting (Lucati and López-Baucells, 2017).
These conditions are generally considered rare in wild populations (Abreu et al. 2013), as they are often associated with various pathologies, including visual (Pérez-Carpinell et al. 1992; Grant et al. 2001; Garipis and Hoffmann 2003) and immunological defects (Carretero et al. 2009; Summers 2009). Hypopigmentation may increase an individual’s conspicuousness, thereby raising susceptibility to predation (Krecsák 2008). Conversely, in some cases, it may reduce predation risk if predators exhibit neophobia, or fear novel stimuli (Mappes et al. 2005)
Hypopigmentation conditions have been documented across all major vertebrate groups in the Neotropics (Abreu et al. 2013), including fish (e.g., Wakida-Kusunoki et al. 2022; Paschoal et al. 2024), amphibians (e.g., Sanabria et al. 2010; Valdez-Villavicencio and Peralta-García 2014), reptiles (e.g., Silva et al. 2010; de Noronha et al. 2013; Sosa-Cornejo et al. 2022), birds (e.g., Ayala-Pérez et al. 2013; Reséndiz-Cruz and Caballero-Jiménez 2016; Palacios-Vázquez 2016; Salgado-Flores and Rodríguez-Ruiz 2022), and mammals (e.g., López-González 2011; Camargo et al. 2014; Arriaga-Flores et al. 2016; Lucati and López‐Baucells 2017; Ramos-Luna et al. 2022). In Mexico, several cases have been reported in bats (see Uieda 2000), including albinism in the common vampire bat (Desmodus rotundus, Uieda 2001; Ramírez et al. 2010), piebaldism in the common fruit bat (Artibeus jamaicensis, Sánchez-Hernández et al. 2010), and leucism in Waterhouse’s leaf-nosed bat (Macrotus waterhousii), the free-tailed bat (Tadarida brasiliensis), the common fruit bat (A. jamaicensis, Sánchez-Hernández et al. 2012), and the ghost-faced bat (Mormoops megalophylla, Hernández-Aguilar and Santos-Moreno 2018). Other documented cases in carnivores include the coyote (Canis latrans, López-González 2011) and the neotropical otter (Lontra longicaudis annectens, Arriaga-Flores et al. 2016), as well as in primates such as howler monkeys (Ramos-Luna et al. 2022) and marine mammals like dolphins (Ortega-Ortiz et al. 2022), to name a few.
In opossums (order Didelphimorphia), hypopig-0mentation has been reported in Brazil (Didelphis marsupialis, Abreu et al. 2013), Colombia (D. marsupialis, Hoyos et al. 2020), Panama (Caluromys derbianus, Fuentes et al. 2024), and Mexico. In Mexico, cases have only been documented on the Yucatán Peninsula, including an albino D. virginiana (Cuxim-Koyoc et al. 2020) and a case of flavism (characterized by cinnamon-colored fur in Didelphis sp.; Tenorio-Rodríguez et al. 2024). This is particularly notable given that opossums (genus Didelphis, Linnaeus, 1758) are widespread and relatively common across Mexico, occurring in nearly all states except for the Baja California Peninsula and some arid regions bordering the United States and the Central Plateau (Ceballos et al. 2002; Gardner and Sunquist 2003). On the Yucatán Peninsula, Didelphis is represented by two sympatric species, D. marsupialis and D. virginiana, with populations of D. virginiana appearing to dominate the northern region over D. marsupialis (Jones et al. 1974; Ruiz-Piña and Cruz-Reyes 2002).
Populations of D. virginiana in the Peninsula may correspond to the subspecies D. v. yucatanensis (Gardner 1973; MacManus 1974), which is reportedly smaller than D. v. californica, although both subspecies share the same coloration pattern (Gardner 1973): dichromatic, with a common dark phase characterized by black legs, feet, and ears; extensive black pigmentation on the tail; dark body coloration extending forward over the top of the head forming a wedge between the eyes; darker sides of the head and neck; and a prominent ocular stripe extending from in front of the eye to a pale spot at the base of the ear, distinctly outlining the white cheek area. If local populations are small, isolated, and subject to inbreeding, we might expect an increased frequency of developmental abnormalities, including coloration disorders (MacManus 1974).
Here, we presented the first documented case of leucism in Didelphis virginiana and the third recorded case of hypopigmentation for this species in the Yucatán Peninsula.
We conducted a photo-trapping survey targeting medium- to large-sized mammals associated with the wildlife crossing structures (pipes, box culverts, and wildlife underpasses) along the Nuevo Xcan–Playa del Carmen Highway, located in the municipalities of Solidaridad and Lázaro Cárdenas, Quintana Roo, Mexico. The region is predominantly flat, with an elevation ranging from 5 to 10 meters above sea level. The climate is warm and sub-humid, with annual mean temperatures ranging from 26°C to ٣٣°C, and an average annual precipitation of approximately ١,٣٠٠ mm, concentrated primarily between June and October (INEGI 2017). The natural vegetation in the area consists of subperennial evergreen forest at various stages of succession (Rzedowski 2006).
We set up 28 monitoring stations that continuously operated from July 2016 to July 2017. Cameras were placed approximately 50cm above the ground in the middle section inside the crossing structure, perpendicular to the entrance aimed at photographing all passing animals (González-Gallina et al. 2018). We considered a camera night a 24-hour period during which the camera was operating. Sampling effort per station was obtained by counting the days the camera was active subtracting days when the camera was not functioning and total sampling effort as the added number of camera nights for each station (Ramesh and Downs 2015).
As part of the project, we also recorded opportunistic observations of road-killed animals encountered while traveling between photo-trapping stations by vehicle. When the condition of the carcasses permitted, individuals were identified to species level using specialized field guides (Reid 2009). To differentiate Didelphis virginiana from D. marsupialis, we relied on diagnostic features such as cheek coloration, whisker color on the muzzle and cheeks, tail length, and the proportion of the tail covered in dark pigmentation. For each observation, we recorded the GPS location and took reference photographs. In the case of photo-trapping data, individuals of the genus Didelphis (D. marsupialis and D. virginiana) were grouped into a single category, as distinguishing between the two species was not feasible in most photographs.
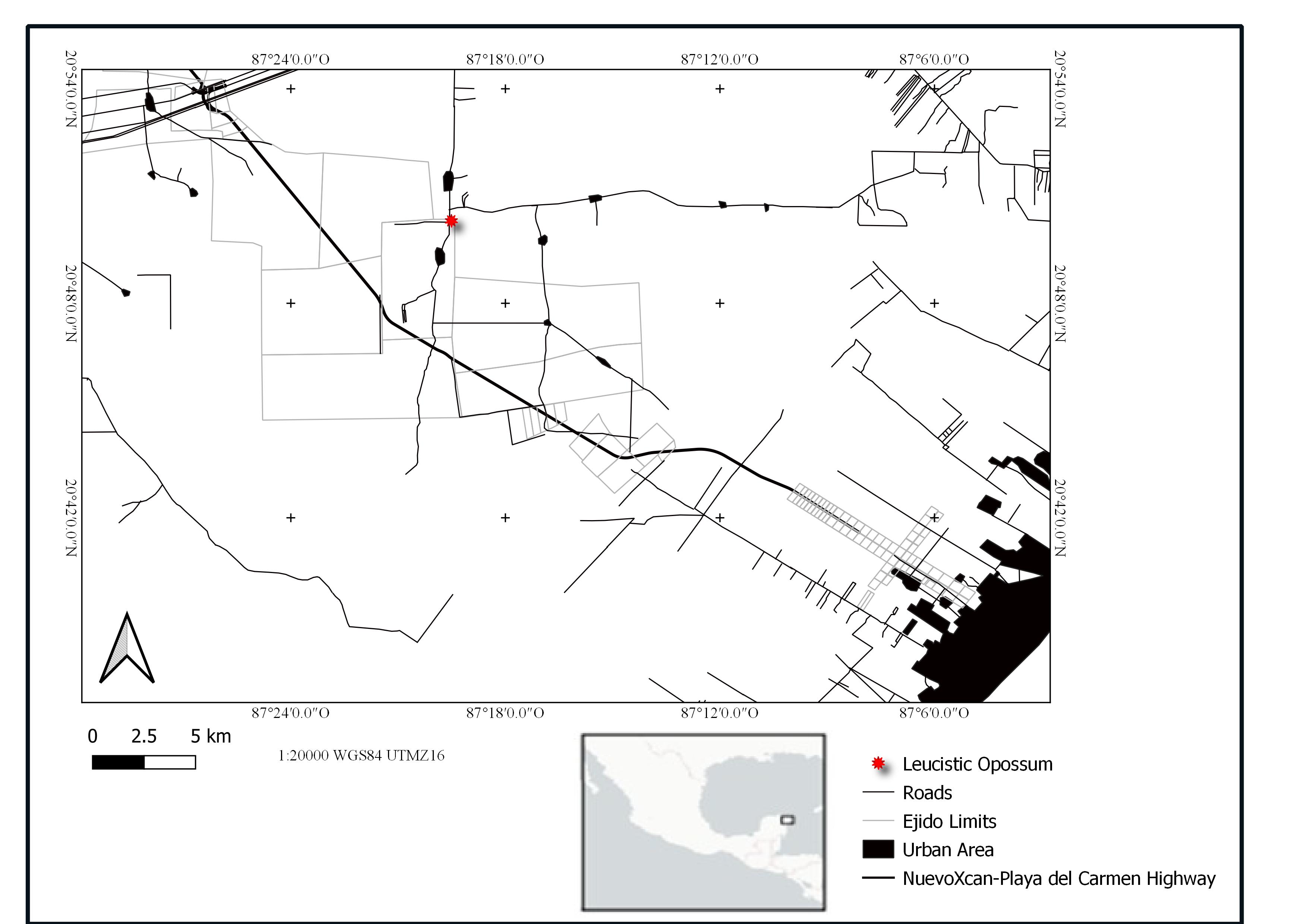
On March 11, 2017, at 15:44 h, we documented a road-killed adult male opossum (Didelphis virginiana) with leucistic coloration along the highway connecting the Mérida–Cancún freeway with the town of Juárez, between the villages of Agua Azul and Juárez (20°50’18.36” N / 87°19’30.75” W) (Figure 1). While traveling this route, we encountered the specimen and examined it for diagnostic characteristics to distinguish between Didelphis species. These included uniformly white whiskers and a tail shorter than the body length, with the dark portion of the tail longer than the white portion—though in this individual, the contrast was less conspicuous (Figure 2)—allowing for identification as D. virginiana. The specimen appeared relatively fresh and based on the degree of rigor mortis and overall condition, we estimated that it had likely died during the night of March 10 because of a vehicle collision. This was the only individual exhibiting abnormal coloration that we encountered during our entire photo-trapping survey in the area.
In addition to this observation, during the survey, we recorded other roadkill specimens, including two opossums (Didelphis marsupialis), two gray foxes (Urocyon cinereoargenteus), two coatis (Nasua narica), and one Yucatán squirrel (Sciurus yucatanensis).
Throughout 10,166 camera trap nights, we obtained 33 records of Didelphis spp. using underpass structures to cross the highway. Of these, nine were inside wildlife-specific crossing structures, 19 utilized box culverts, and five employed concrete pipes.
Pigmentation anomalies in natural populations of Neotropical mammals have been considered rare (Abreu et al. 2013). To provide local context, from November 2012 to July 2013, the consultancy SEGA S.A. de C.V. conducted biological monitoring along the Nuevo Xcan–Playa del Carmen highway (Hidalgo-Mihart et al. 2013). This effort involved 54 camera traps across 81 photo-trapping stations, resulting in a total of 7,937 trap-nights. Opossums (Didelphis spp., as the species are difficult to distinguish in photographs) emerged as a dominant species, with 325 independent records. Also, González-Gallina et al. (2018) reported an additional 36 records of Didelphis. Across both monitoring efforts—361 total records—only a single case of color abnormality was detected within the genus. This single record of a pigmentation anomaly through roadkill rather than photo-trapping shows that opportunistic highway surveys can yield important complementary records (González-Gallina et al. 2016).
Hypopigmented animals are generally more susceptible to fitness declines compared to hyperpigmented (melanic) individuals. The occurrence of hypopigmented wild mammals appears to be more frequent in areas with high levels of human activity, where fragmented populations are more prone to environmental stress and inbreeding (Guestalla et al. 2021; Cotts et al. 2024). This pattern may reflect underlying environmental conditions in the Yucatán Peninsula, as all reported cases of hypopigmentation in Didelphis from Mexico originate from this region. However, in our study area, low population, isolation and inbreeding are unlikely explanations, as Didelphis (including both virginiana and marsupialis) was the most abundant species recorded (Hidalgo-Mihart et al. 2013). Still, these genetic disorders are naturally occurring (sometimes by random mutations) but rare (Abreu et al. 2013) as in this case this record represents 0.28 of all records.
If white pelage is better explained by background matching (Caro 2009), we propose that the light-colored karstic substrate that characterizes much of the Yucatán Peninsula (Bautista et al. 2015) may play a role in the survival of these hypopigmented individuals. The local soil type, known as sascab (meaning “white dirt” in Maya), could favor them by reducing their detectability to predators, thus mitigating the fitness costs often associated with conspicuous coloration. In this context, the cryptic advantage provided by sascab may reduce the non-physiological (e.g., predation-related) adverse effects of hypopigmentation observed in other environments.
This could help explain how the albino opossum reported by Cuxim-Koyoc et al. (2020) reached adulthood despite likely visual impairments—limitations not typically present in leucistic individuals such as the one we report here. It might also account for our observation of a leucistic female great curassow (Crax rubra) captured during photo trapping in the same locality (Hidalgo-Mihart et al. 2013).
We suspect that color patterns in mammals may be influenced by both habitat characteristics and human-induced factors (Ausband and Krohner 2022). Further research is necessary to better understand how selective pressures related to habitat, life history, developmental stage, and season contribute to the evolutionary mechanisms underlying external coloration. To advance this understanding, it is necessary to compile enough records of chromatic disorders across a wide range of species and taxonomic groups worldwide. This would allow us to begin identifying potential environmental patterns and, importantly, to determine which variables may promote the occurrence of such chromatic anomalies in wild populations.
Acknowledgement
For funding this project, CONAHCYT for the doctoral grant 335814/232663 given to A. González Gallina together with Conservación Panthera México A.C. and the Rufford Foundation for the grant number 22395-1.
Literature cited
Abreu, M. S. L., et al. 2013. Anomalous color in Neotropical mammals: a review with new records for Didelphis sp. (Didelphidae, Didelphimorphia) and Arctocephalus australis (Otariidae, Carnivora). Brazilian Journal of Biology 73:185-194.
Acevedo, J., and M. Aguayo. 2008. Leucistic South American sea lion in Chile, with a review of anomalously color in otariids. Revista de Biología Marina y Oceanográfica ٤٣:٤١٣–٤١٧.
Arriaga-Flores, J. C., et al. 2016. Leucism in neotropical otters (Lontra longicaudis annectens) from México. The Southwestern Naturalist 61:63-68.
Ausband, D. E., and J. M. Krohner. 2022. Carnivores in color: pelt color patterns among carnivores in Idaho. Journal of Mammalogy 103:598-607.
Ayala-Pérez, V., et al. 2013. Observations of leucistic shorebirds in NW México. Wader Study Group Bulletin 120:159-161.
Bautista, F., et al. 2015. Actualización del mapa de suelos del Estado de Yucatán México: Enfoque geomorfopedológico y WRB. Ecosistemas y Recursos Agropecuarios ٢:٣٠٣-٣١٥.
Camargo, I., et al. ٢٠١٤. First record of leucism in the genus Peromyscus (Mammalia: Rodentia). Western North American Naturalist 74:366-368.
Caro, T. 2005. The adaptive significance of coloration in mammals. BioScience 55:125-136.
Caro, T. 2009. Contrasting coloration in terrestrial mammals. Philosophical Transactions of the Royal Society B. Biological Sciences 364:537-548.
Carretero, P., et al. 2009. Síndrome de Griscelli-Prunieras: a propósito de dos casos. Anales de Pediatría ٧٠:١٦٤-١٦٧.
Ceballos, G., et al. ٢٠٠٥. Diversidad y Conservación de los Mamíferos de México. Pp. ٢١-٦٦. en: Los Mamíferos Silvestres de México (G. Ceballos y G. Oliva, coords.) Conabio-Fondo de Cultura Económica. México D.F.
Cotts, L. et al. 2024. Unveiling the Colors of Mustelids: A Historical Review on the Emergence of Chromatic Disorders and Their Ecological Effects in Mustelids Worldwide with Report of the First Case of Erythrism in Eira Barbara (Carnivora, Mustelidae). Animals 14:354
Cuxim-Koyoc, A., et al. 2020. Albinism in Didelphis virginiana (Kerr, 1792): the first reported case in México. Mammalia 84:144-149.
Delhey, K. 2019. A review of Gloger’s rule, an ecogeographical rule of colour: Definitions, interpretations and evidence. Biological Reviews 94:1294-1316.
de Noronha, J. D. C., et al. 2013. Record of leucism in Pseudoboa nigra (Serpents: Dipsadidae) in southern Amazon, Brazil. Herpetology Notes 6:81-82.
Fertl, D., and P. Rosel. 2002. Albinism. Pp. 16-18 in Encyclopedia of Marine Mammals (Perrin W. F., B. Würsig, and J.G.M. Thewissen, eds.). Academic Press. San Diego, EE.UU.
Fertl, D., et al. 2004. An update on anomalously white cetaceans, including the first account for the pantropical spotted dolphin (Stenella attenuata graffmani). Latin American Journal of Aquatic Mammals 3:163–166.
Fuentes, R., et al. 2024. Report of coloration anomalies in mammals from Panama. Neotropical Biology and Conservation 19:333-345.
Gardner, A. 1973. The Systematics of the Genus Didelphis (Marsupialia: Didelphidae) in North and Middle America. Special Publication, The Museum, Texas Tech University. Vol. 4.
Gardner, A., and M. E. Sunquist. 2003. Opossum, Didelphis virginiana. Pp. ٣–٢٩ in Wild mammals of North America: biology, management, and conservation (Feldhamer, G. A., A. B. C. Thompson, and J. A. Chapman, eds.). John Hopkins University Press. Baltimore, EE.UU.
Garipis, N., and K. P. Hoffmann. 2003. Visual field defects in albino ferrets (Mustela putorius furo). Vision research 43:793-800.
González-Gallina, A., et al. 2016. Roadkills as a complementary information source for biological surveys using rodents as a model. Journal of Mammalogy 97:145-154.
González-Gallina, A., M. G. Hidalgo-Mihart, and V. Castelazo-Calva. 2018. Conservation implications for jaguars and other neotropical mammals using highway underpasses. PLoS One 13:e0206614.
Grant, S., et al. 2001. Rod photopigment deficits in albinos are specific to mammals and arise during retinal development. Visual neuroscience 18:245-251.
Guastalla, M.G., et al. 2021. The mysterious white deer: Anomalous coloring in different Neotropical deer. Mammalian Biology 101:665–673.
Hernández-Aguilar, I., and A. Santos-Moreno. 2018. First records of hypopigmentation disorders in the Peters’ ghost-faced bat Mormoops megalophylla (Chiroptera, Mormoopidae). Mammalia ٨٢:٦١٨-٦٢١.
Hidalgo-Mihart, M. G., et al. ٢٠١٣. Monitoreo de mamíferos del proyecto carretero Ramales Cedral-Tintal y Tintal-Playa del Carmen con una longitud de ٥٤ km, en el estado de Quintana Roo. Informe Final. Servicios Estratégicos para la Gestión Ambiental S. A. de C.V. Available through SEGA Planeación y Desarrollo (segapd.com).
Hoyos, R., et al. ٢٠٢٠. Primer registro de leucismo en Didelphis marsupialis Linnaeus, ١٧٥٨ (Didelphimorphia: Didelphidae) en Colombia. Galemys ٣٢:٦٤-٦٨.
Instituto Nacional de Estadística y Geografía (INEGI). 2017. In: http://www.beta.inegi.org. mx/app/biblioteca/ficha.html?upc=702825097912. Accessed on March 2018.
Jones, J. K. Jr., H. H. Genoways, and J. D. Smith. 1974. Annotated checklist of mammals of the Yucatan Peninsula, Mexico. III. Marsupialia, Insectivora, Primates, Edentata, Lagomorpha. Occasional Papers of the Museum, Texas Tech University 23:1-12.
Krecsák, L. 2008. Albinism and leucism among European Viperinae: a review. Russian journal of Herpetology 15:97-102.
Lamoreux, M.L. et al. 2010. The Colors of Mice. A Model Genetic Network. Wiley-Blackwell, Bryan, Texas, USA.
López-González, C. A. ٢٠١١. Registro de un coyote (Canis latrans) blanco en el desierto chihuahuense de Durango, México. Acta Zoológica Mexicana (Nueva Serie) ٢٧:٨٧١–٨٧٣.
Lucati, F. and A. López‐Baucells. 2017. Chromatic disorders in bats: a review of pigmentation anomalies and the misuse of terms to describe them. Mammal Review 47:112-123.
McManus, J. J. 1974. Didelphis virginiana. Mammalian species (40):1-6.
Mappes, J., N. Marples, and J.A. Endler 2005. The complex business of survival by aposematism. Trends in Ecological Evolution 20:598–603.
Miller, J. D. 2005. All about albinism. Missouri Conservationist 66:5–7.
Ortega-Ortiz, C. D., et al. 2022. Observations of Hypopigmented Dolphins Sighted in Mexican and Alaskan Waters (2012-2013). Aquatic Mammals ٤٨:٧٤٦-٧٥٢.
Palacios-Vázquez, A. J. ٢٠١٦. Primer registro de leucismo total en el tirano tijereta rosado (Tyrannus forficatus) en México. Huitzil 17:229-233.
Paschoal, L. R. P., et al. 2024. Leucism in the marbled swamp eel Synbranchus marmoratus Bloch, 1795 (Synbranchiformes: Synbranchidae). Biota Neotropica 24:e20231592.
Pérez-Carpinell, J., et al. 1992. Vision defects in albinism. Optometry and Vision Science 69:623-628.
Ramesh T, and C.T. Downs. 2015. Impact of land use on occupancy and abundance of terrestrial mammals in the Drakensberg Midlands, South Africa. Journal of Nature Conservation 23:9–18.
Ramos-Luna, J., et al. 2022. Anomalous pigmentation in American primates: Review and first record of a leucistic black howler monkey in Southeast México. Folia Primatologica ٩٣:87-95.
Reid, F. A. 2009. A field guide to the Mammals of Central America and Southeast Mexico. Second Edition. Oxford University Press. New York, EE.UU.
Reséndiz-Cruz, I., and R. Caballero-Jiménez. 2016. Primer registro de leucismo parcial en el mirlo pardo (Turdus grayi) para México. Huitzil 17:225-228.
Ruiz-Piña, H. A., and A. Cruz-Reyes. 2002. The opossum Didelphis virginiana as a synanthropic reservoir of Trypanosoma cruzi in Dzidzilché, Yucatán, México. Memorias do Instituto Oswaldo Cruz 97:613-620.
Rzedowski, J. 2006. Vegetación de Mexico. 1ra. Edición digital. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. México.
Salgado-Flores, J. L., and E. R. Rodríguez-Ruiz. ٢٠٢٢. Leucismo parcial en el zanate mayor (Quiscalus mexicanus) en México. Huitzil ٢٣:١-٤.
Sanabria, E. A., L. B. Quiroga, and A. Laspiur. 2010. First Record of Partial Albinism and Scoliosis in Odontophrynus occidentalis Tadpoles (Anura: Cycloramphidae). Brazilian Archives of Biology and Technology 53:641-642.
Sánchez-Hernández, C., et al. 2010. Five albino bats from Guerrero and Colima, Mexico. Chiroptera Neotropical 16:522–527.
Sánchez-Hernández, C., et al. 2012. Leucism in five species of bats from Mexico. Chiroptera Neotropical 18:1123-1127.
Silva, F., C. Assis, and F. M. Quintela. 2010. Albinism in a Liophis miliaris (Linnaeus, 1758) (Serpentes: Dipsadidae) from Minas Gerais state, southern Brazil. Herpetology Notes 3:171–172.
Sosa-Cornejo, I., et al. 2022. Leucism: the prevalent congenital malformation in the olive ridley sea turtle of northwestern México. Diseases of Aquatic Organisms 152:61-71.
Summers, C. G. 2009. Albinism: classification, clinical characteristics, and recent findings. Optometry and Vision Science 86:659-662.
Tenorio-Rodríguez, P. A., L. Cab-Sulub, and L. D. Moreno-Figueroa. 2024. First record of cinnamon coloration in opossum Didelphis sp. in México. Therya Notes ٥:192-196.
Uieda, W. 2000. A review of complete albinism in bats with five new cases from Brazil. Acta Chiropterolo-gica ٢:97-105.
Uieda, W. 2001. Behavior of an albino vampire bat, Desmodus rotundus (E. Geoffroy) (Chiroptera, Phyllostomidae), in captivity. Revista Brasileira de Zoologia 18:641-644.
Valdez-Villavicencio, J. H., and A. Peralta-García. 2014. Aneides lugubris (arboreal salamander): leucism. Herpetological Review ٤٥:108.
Wakida-Kusunoki, A. T., V. Anislado-Tolentino, and L. F. D. Moral-Flores. 2022. First report of synophthalmia and leucism in a bull shark (Carcharhinus leucas) embryo from the southeastern Gulf of México. Latin American Journal of Aquatic Research 50:760-766.
Walter, H. E. 1914. Genetics: An introduction to the study of heredity. Macmillan. Michigan, EE.UU.
Zalapa, S., et al. 2016. Coloración atípica en murciélagos: frecuencia y fenotipos en Norte y Centroamérica e islas del Caribe y nuevos casos para México y Costa Rica. Revista Mexicana de Biodiversidad 87:474-482.
Associated editor: Beatríz Bolívar Cimé.
Submitted: January 14, 2025; Reviewed: April 23, 2025.
Accepted: July 6, 2025; Published on line: August 12, 2025.