THERYA NOTES 2024, Vol. 5 : 249-254 DOI: 10.12933/therya_notes-24-181 ISSN 2954-3614
Observations of feeding attempts on Baird’s tapirs (Tapirella bairdii) by common vampire bats (Desmodus rotundus) in Corcovado National Park, Costa Rica
Observaciones de intentos de alimentación en la danta centroamericana (Tapirella bairdii) por parte de murciélagos vampiros comunes (Desmodus rotundus) en el Parque Nacional Corcovado, Costa Rica
April Shipley-Mellon1, Alejandro Azofeifa2, Danny Herrera2, Evelyn T. Solano-Mora2, Guido Saborío-R2, and Erik R. Olson1*
1Department of Natural Sciences, Northland College, Ashland, WI 54806, U.S.A. E-mail: shipleymellonapril@gmail.com (AS-M); eolson@northland.edu (ERO).
2Área de Conservación Osa, Sistema Nacional de Áreas de Conservación. Oficinas Regionales ACOSA. 800 mts Norte de Pista de aterrizaje de Golfito, C. P. 60701, Golfito. Puntarenas, Costa Rica. E-mail: alejandro.azofeifa@sinac.go.cr (AA); danny.herrera@sinac.go.cr (DH); tatiana.solano@sinac.go.cr (ETS-M); guido.saborio@sinac.go.cr (GS-R).
*Corresponding author
Amit and Valverde-Zúñiga reported the first observations of common vampire bats (Desmodus rotundus) attempting to feed on Baird’s tapirs (Tapirella bairdii) in Costa Rica. They reported behaviors they believed to be consistent with anti-parasitism behaviors for the species. We provide additional evidence validating their observations, making the case that such anti-parasitism behaviors may have been selected for and may be widespread within neotropical mammals. As part of a long-term wildlife monitoring and conservation effort, we maintained a camera trap network within Corcovado National Park, Costa Rica since 2015. Our camera trap network consisted of 13 to 56 paired or unpaired cameras placed in a 4 x 4 km grid. In 2021, we captured video footage of one D. rotundus crawling up to a T. bairdii in an apparent feeding attempt as a second D. rotundus appeared to be waiting nearby. The feeding attempt was evaded due to what appears to be behaviors consistent across species, time, and space, suggestive of an evolved behavioral response to feeding attempts by D. rotundus. In 2023, we documented a second event of an apparent feeding attempt by a single D. rotundus on a juvenile T. bairdii walking through our camera’s detection area. Our observation combined with the observations of others suggests that a sudden movement that forces D. rotundus off the body may be an effective strategy for preventing an attack. Some neotropcial wild mammals may have adapted specific avoidance strategies for dealing with D. rotundus.
Key words: Camera trap; defensive behavior; feeding attempt; group-feeding; interspecific interaction; rabies virus.
Amit y Valverde-Zúñiga reportaron las primeras observaciones de Desmodus rotundus intentando alimentarse de Tapirella bairdii en Costa Rica. Ellos reportaron comportamientos que creían consistentes con comportamientos de antiparasitismo para las dantas. Proporcionamos evidencia adicional que argumenta que tales comportamientos antiparasitarios pueden haber sido seleccionados y estar extendidos entre los mamíferos neotropicales. Mantuvimos una red de cámaras trampa dentro del Parque Nacional Corcovado, Costa Rica desde 2015. Nuestra red consistió en 13 a 56 cámaras emparejadas o no emparejadas, colocadas en una cuadrícula de 4 x 4 km. En 2021 capturamos vídeo de un D. rotundus arrastrándose hasta un T. bairdii en un aparente intento de alimentación, mientras un segundo D. rotundus parece estar esperando cerca, mostrando una táctica de alimentación en grupo bien conocida de esta especie. Sin embargo, la danta evadió el intento de alimentación debido a lo que parece ser un comportamiento consistente a través de algunas especies en el tiempo y el espacio. Esto sugiere una respuesta conductual evolutiva a los intentos de alimentación por parte de D. rotundus. En 2023, documentamos un segundo evento entre un individuo de D. rotundus y un T. bairdii juvenil que caminaba por el área de detección de nuestra cámara. Nuestras observaciones, combinadas con las observaciones de otros, sugieren un movimiento repentino que obliga a D. rotundus a apartarse del cuerpo de la danta, lo cual es una estrategia eficaz para evitar un ataque.
Palabras clave: Alimentación grupal; cámara trampa; comportamiento de defensa; intento de alimentación; interacción interespecífica; virus de la rabia.
© 2024 Asociación Mexicana de Mastozoología, www.mastozoologiamexicana.org
Interactions between parasitic vampire bats (family Desmodontidae) and their hosts are only recently being understood. Amit and Valverde-Zúñiga (2022) recently documented the first observations of interactions between the common vampire bat (Desmodus rotundus) and Baird’s tapir (Tapirella bairdii, also referred to as Tapirus bairdii). They observed T. bairdii behaviors such as bucking, shaking, charging, running, and spinning to avert D. rotundus feeding attempts. Such observations alone provide insights into how potential hosts avoid feeding attempts by vampire bats. Taken independently, these observations are interesting, but when examined within the broader literature on the topic we may begin to gain further insights into the prevalence of such behaviors. For example, Gnocchi and Srbek-Araujo (2017) reported some of the same behaviors for lowland tapirs (Tapirus terrestris) of the Amazon. Herein, we contribute additional observations to this growing body of literature regarding D. rotundus feeding attempts on wild and domestic prey, and we draw comparisons amongst the published observations to suggest an evolved behavioral response to D. rotundus feeding attempts that spans species, space, and time.
Desmodus rotundus is a hematophagous bat species (Turner 1975), with its preferred prey being medium-to-large-sized mammals (Gnocchi and Srbek-Araujo 2017). While mainly feeding on livestock due to abundance and availability (Galetti et al. 2016), D. rotundus also feeds on other domestic animals and both native and non-native wildlife (Gnocchi and Srbek-Araujo 2017). Attacks on wildlife are typically documented via bite scars on prey (Sánchez-Cordero et al. 2011; Castellanos and Banegas 2015) which are characterized by the sharp incisors of the species (Arellano-Sota 1988). However, some researchers have opportunistically documented feeding attempts via camera traps.
During a feeding event, D. rotundus consume between 50 % and 100 % of their body weight in blood (Wilkinson 1990). If D. rotundus does not feed within 50 to 60 hr after a prior feeding, the individual will not have enough energy to maintain proper body temperature, resulting in death (Carter and Wilkinson 2013). Thus, D. rotundus cannot go longer than 2 nights without feeding (Wilkinson 1990; Carter and Wilkinson 2013). To improve feeding and survival success, D. rotundus are known to return to the same site to feed multiple nights in a row or to share feeding sites (Gnocchi and Srbek-Araujo 2017). Thus, the blood loss impacts of D. rotundus parasitism may be considerable for some species (i.e., smaller bodied species) and are generally poorly understood.
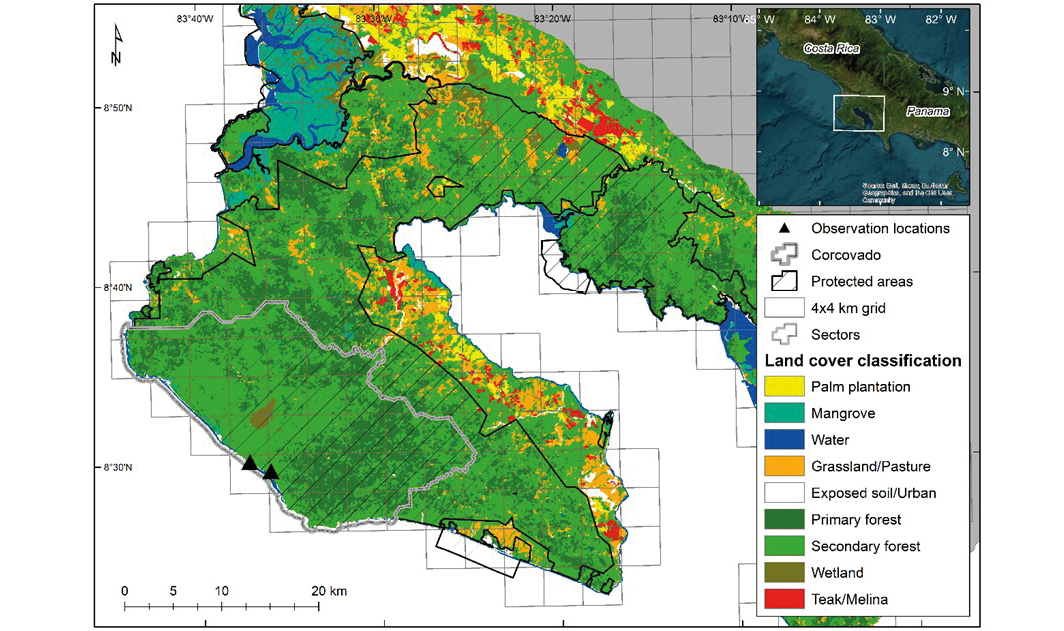
As part of a long-term wildlife monitoring and conservation effort (i.e., JaguarOsa project), we have implemented and maintained a camera trap network within Corcovado National Park, Costa Rica since 2015 (Olson et al. 2022; see also Olson et al. in press). Corcovado National Park is one of Costa Rica’s largest national parks (424 km2; Figure 1; Olson et al. 2022). It consists of a sizeable and globally rare lowland tropical forest surrounded by a mountainous evergreen tropical forest. These habitats support a plethora of species, including the globally endangered T. bairdii (García et al. 2016; Olson et al. 2022). Considered a core habitat area for T. bairdii, Corcovado is estimated to support a density of 0.81 tapirs/km2 (Schank et al. 2017). Our camera trap network consisted of 13 to 56 paired or unpaired (i.e., 1 or 2 cameras, either Reconyx, Bushnell, or UOVision camera traps) camera stations depending on survey year, with an average survey effort of 1,453 trap nights from 2015 to 2021 (Olson et al. 2022). Historically, our surveys were mainly run from February to June or July and cameras with photo or video capability are placed within a 4 x 4 km wildlife monitoring grid (see Olson et al. 2022 and Olson et al. in press for detailed summaries of survey efforts). We installed cameras to cover between 23 % and 59 % of grid cells within the park, depending on the survey year (Olson et al. 2022; Figure 1). In 2021, 2022, and 2023, we ran surveys from January to December with a total of 34, 56, and 45 camera stations, respectively.
On March 7, 2021 at 01:20 hr near Corcovado Beach (8° 30' 03.6" N, 83° 37' 15.6" W, we observed an adult female T. bairdii (Figure 2) resting on the ground when one D. rotundus crawled towards it on the forest floor and contacted the right posterior side of the body in an apparent effort to make a feeding attempt (Figure 2; see also Galetti et al. 2016). The T. bairdii made a sudden movement and stood up, spun, and took an aggressive stance (front feet spread more than shoulder width apart and bent at the elbow). These actions resulted in the D. rotundus retreating and an unsuccessful feeding attempt. The female T. bairdii then investigated (i.e., smelled) the area where the D. rotundus made contact. After a while, 2 D. rotundus individuals flew through the frame causing the T. bairdii to spin and investigate. The 2 D. rotundus are then seen following the T. bairdii as it runs away and out of the camera frame. In November 2023 at 03:56 hr, near Laguna Cocovado (8° 29' 34.8" N, 83° 36' 10.8” W) we documented a second event of an adult female tapir walking through our camera’s detection area followed closely by a juvenile. An individual D. rotundus closely followed the juvenile tapir as it walked through the detection area first on the ground and then in flight, in what we believe was a clear attempt to parasitize the juvenile (Figure 3). However, in this observation, both the adult and juvenile tapirs seem to be unaware or unbothered by the presence of the bat (see Appendix 1 for full video of both events). Of the 1,891 events of T. bairdii (i.e., 30 min window of time with a T. bairdii detection) between 2015 and 2023 (~77 % of which have occurred between 18:00 and 06:00 hr), these are the only observations we have of such an interaction (~ 0.14 % of nighttime events). We have observed at least 2 additional events with T. bairdii and flying bats but are unable to confirm the species of bat. It is possible that we failed to detect additional D. rotundus events due to either poor quality imagery or the cryptic nature and small size of the species.
The observations by Amit and Valverde-Zúñiga (2022) document behaviors such as shaking, running, charging, and spinning as a defense utilized by T. bairdii to prevent a successful feeding attempt by D. rotundus. A similar movement in response to feeding attempts by D. rotundus has been documented with 2 other species via camera traps in Brazil; Tapirus terrestris (Gnocchi and Srbek-Araujo 2017) and Mazama temama (Galetti et al. 2016). Both incidents resulted in failed feeding attempts for D. rotundus. These observations are similar to ours and those of Amit and Valverde-Zúñiga (2022) and help to validate these behaviors as an effective strategy for preventing an attack by D. rotundus. Our additional observation of D. rotundus attempting to feed on a juvenile tapir may indicate that these behaviors are not yet obvious to juveniles. It is possible that D. rotundus may have more success feeding on naïve juveniles compared to adults but given that this is the only documented interaction so far, we are limited in our understanding.
Galetti et al. (2016) also report that D. rotundus has been observed chasing adult capybaras (Hydrochoerus hydrochaeris). These observations across species, space, and time, suggest that some neotropical mammals have adapted specific avoidance strategies for dealing with D. rotundus. Compared to wild mammals, we know that domestic mammals generally have reduced anti-predator responses- likely as a result of domestication for less risky behaviors for human handlers (Geffroy et al. 2020). The loss of stress responses and other behaviors appears to make domestic mammals more vulnerable to wild predators (Kaiser et al. 2015; Geffroy et al. 2020; Solberg et al. 2020). The same seems plausible for behaviors that could result in failed feeding attempts or even injury or mortality for D. rotundus. Such a difference could also help explain why D. rotundus tends to prefer domestic mammals over wild mammals in sites where both are present (Amit and Valverde-Zúñiga 2022). Our observations also suggest that juvenile wild neotropical mammals may be more vulnerable than adults to D. rotundus parasitism; however, further research is required.
The main risk associated with interactions between D. rotundus and wildlife is the potential for transmission of dangerous pathogens, particularly rabies virus (RABV), a viral zoonotic disease that attacks the central nervous system (Fisher et al. 2018) and is known to be carried by D. rotundus (Gnocchi and Srbek-Araujo 2017; León et al. 2021). While feral and domestic dogs are typically the main transmitter of RABV across Central America (León et al. 2021), recent efforts by the government of Costa Rica have eliminated canine rabies outbreaks. Now, D. rotundus serves at the main reservoir of RABV across Costa Rica (León et al. 2021). RABV vaccinations remain an effective tool for preventing outbreaks and death for livestock and humans (Gnocchi and Srbek-Araujo 2017), but mitigation of rabies outbreaks in the wild is far more complicated. Using their estimate of a T. terrestris’ chance of being attacked by D. rotundus (0.11) and D. rotundus–RABV prevalence of 0.014, Galetti et al. (2016) estimated the probability of rabies transfer to T. terrestris in the Pantanal at 0.15 %. While we are not able to assess the probability of RABV transfer to T. bairdii, we suspect the probability is much lower because we have only detected 2 interactions of many T. bairdii events (2 of at least 1,456 events). Furthermore, we suspect that D. rotundus is found at lower densities in Costa Rica compared to Brazil because of Brazil’s larger domestic livestock populations (O’Donoghue et al. 2019). RABV is well-studied in human and domestic animal populations, but studies regarding RABV in the wild are still lacking. Our observations may provide insights into potential RABV transmission for wild neotropical mammals in this region of Costa Rica and may inspire researchers and practitioners in the One Health field to further explore the importance of such findings.
Our observations confirm that D. rotundus feeds, or at least attempts to feed, on T. bairdii in the wild. When put within the context of the broader published behavioral observations, failed D. rotundus feeding attempts on wild mammals suggest that defensive behaviors such as those reported by Galetti et al. (2016), Gnocchi and Srbek-Araujo (2017), and Amit and Valverde-Zúñiga (2022), i.e., bucking, charging, spinning, running, shaking, and even kicking and biting – may reduce successful parasitism attempts by D. rotundus. While the implications of such interactions remain less clear, these similar observations across species, space, and time provide a strong basis for understanding how these cryptic and rare scenarios unfold. We encourage further reporting of such observations and future research into the degree and implications of such interactions.
Acknowledgements
Thank you to the staff and administration of Corcovado National Park and local guides, SINAC-ACOSA, and Northland College for making this research possible. We thank C. Mischler and J. and B. Matzinger for their continued support of this project. We thank numerous undergraduate students for their hard work and assistance in data management for this project. We thank P. Bonk, P. Holt, J. Moore, and M. Wickman for their constant administrative support. We thank the anonymous reviewers for their insightful comments and suggestions on an earlier version of this manuscript. This research is dedicated to the memory of Parker J. Matzinger; he was a burst of light.
Literature cited
Amit, R., and N. Valverde-Zúñiga. 2022. Bucking and charging defense of Baird’s tapir (Tapirella bairdii) from common vampire bats (Desmodus rotundus). Therya Notes 3:147-152.
Arellano-Sota, C. 1988. Biology, ecology, and control of the vampire bat. Reviews of Infectious Diseases 10:S615-S619.
Carter, G. G., and G. S. Wilkinson. 2013. Food sharing in vampire bats: reciprocal help predicts donations more than relatedness or harassment. Proceedings of the Royal Society B 28.
Castellanos, A., and G. Banegas. 2015. Vampire bats bite lowland tapirs in Yasuni National Park, Ecuador. Newsletter of IUCN/SSC Tapir Specialist Group 24:33.
Fisher, C. R., et al. 2018. The spread and evolution of rabies virus: conquering new frontiers. Nature reviews Microbiology 16:241-255.
Galetti, M., et al. 2016. Liquid lunch- vampire bats feed on invasive feral pigs and other ungulates. Frontiers in Ecology and the Environment 14:9.
García, M., et al. 2016. Tapirus bairdii. The IUCN Red List of Threatened Species 2016. www.iucnredlist.org. Accessed on January 10, 2022.
Geffroy, B., et al. 2020. Evolutionary dynamics in the Anthropocene: Life history and intensity of human contact shape antipredator responses. PLOS Biology 18:9.
Gnocchi, A. P., and A. C. Srbek-Araujo. 2017. Common vampire bat (Desmodus rotundus) feeding on lowland tapir (Tapirus terrestris) in an Atlantic Forest remnant in southeastern Brazil. Biota Neotropica 17:3.
Kaiser, S., et al. 2015. Domestication affects the structure, development and stability of biobehavioural profiles. Frontiers in Zoology 12:S19.
León, B., et al. 2021. Rabies in Costa Rica - next steps towards controlling bat-borne rabies after its elimination in dogs. The Yale Journal of Biology and Medicine 94:311-329.
O’Donoghue, E., et al. 2019. USDA agricultural predictions to 2028. Office of the Chief Economist, World Agricultural Outlook Board, U.S. Department of Agriculture. Prepared by the Interagency Agricultural Projections Committee. Long-term Projections Report OCE-2019-1. March 2019. Washington, D. C., U.S.A.
Olson, E. R., et al. 2022. Wildlife monitoring report for Corcovado National Park, Costa Rica-2021. Technical Report. Northland College. Wisconsin, U.S.A.
Olson, E. R., et al. In press. Conservation Crisis? Status of jaguars Panthera onca in Corcovado National Park, Costa Rica. Oryx.
Sánchez-Cordero, V., et al. 2011. Vampire bats, Desmodus rotundus, feeding on white-tailed deer, Odocoileus virginianus. Mammalia 75:91-92.
Schank, C. J., et al. 2017. Using a novel model approach to assess the distribution and conservation status of the endangered Baird’s tapir. Diversity and Distributions 23:1459-1471.
Solberg, M. F., et al. 2020. Domestication leads to increased predation susceptibility. Scientific Reports 10:1929.
Turner, D. C. 1975. The vampire bat; a field study in behavior and ecology. John Hopkins University Press. Baltimore, U.S.A.
Wilkinson, G. S. 1990. Food sharing in vampire bats. Scientific American 262:76-83.
Associated editor: Tamara M. Rioja-Paradela.
Submitted: April 24, 2024; Reviewed: September 18, 2024.
Accepted: October 3, 2024; Published on line: October 14, 2024.
Appendix 1
Video documenting two feeding attempts by common vampire bats, Desmodus rotundus on Baird’s tapirs Tapirella bairdii in Corcovado National Park, Costa Rica, in 2021 and 2023. https://www.youtube.com/watch?v=9refUJASudY.
Figure 1. Location of observations of common vampire bat Desmodus rotundus feeding attempts on Baird’s tapirs Tapirella bairdii in Corcovado National Park, Costa Rica (2015-2023).

Figure 2. a) Frame of a video documenting a common vampire bat Desmodus rotundus crawling up to an adult female Baird’s tapir Tapirella bairdii in an apparent feeding attempt in Corcovado National Park, Costa Rica in 2021. To see the full-length video of the interaction with annotations click here; b) adult female T. bairdii standing up after attempted parasitism by a D. rotundus in Corcovado National Park, Costa Rica 2021.

Figure 3. A frame of a video documenting a common vampire bat Desmodus rotundus following a juvenile Baird’s tapir Tapirella bairdii in an apparent feeding attempt in Corcovado National Park, Costa Rica in 2023. To see the full-length video of the interaction with annotations click here.